Обмен веществ и: Как улучшить обмен веществ в организме — что делать, чтобы наладить метаболизм, продукты нормализующие пищеварение
Жировой обмен веществ связан с продолжительностью жизни
Международная группа ученых продемонстрировала влияние липидов на продолжительность жизни. Ученые исследовали 669 образцов из 6 различных тканей 35 видов млекопитающих и пришли к выводу, что липидный метаболизм оказывает существенное влияние на максимальную продолжительность жизни того или иного вида. Эта работа проливает свет на молекулярные механизмы определения продолжительности жизни и механизмы, лежащие в основе процессов старения.
Результаты исследования опубликованы в журнале Nature Scientific Reports.
Максимальная продолжительность жизни — это стабильная характеристика того или иного биологического вида. Несмотря на то, что за последние пару сотен лет средняя продолжительность жизни значительно возросла, максимальная продолжительность жизни человека не изменилась и составляет приблизительно 120 лет. Если мы хотим жить дольше, необходимо понять, от чего зависит наш максимальный возраст.
Максимальная продолжительность жизни даже среди близкородственных видов млекопитающих может варьировать в десятки раз.
Профессор Сколтеха Филипп Хайтович. Фото: Sk.ru
Также ученые обнаружили, что химическое строение липидов оказывает влияние на продолжительность жизни. Например, такие структурные липиды, как сфинголипиды, входящие в состав клеточной мембраны долгоживущих видов, более насыщены водородом, чем сфинголипиды короткоживущих видов. Ученые объясняют этот феномен большей устойчивостью насыщенных липидов к окислительному стрессу, которому наши клетки подвергаются при старении. В то же время для липидов, участвующих в энергетических процессах, зависимость ровно противоположная. Ненасыщенных энергетических липидов у долгожителей больше, чем у короткоживущих видов. Авторы статьи говорят, что это связано с тем, что такие липиды клеткам сложнее превращать в энергию, а замедленный обмен веществ — одна из характеристик долгоживущих видов.
Ученые объясняют этот феномен большей устойчивостью насыщенных липидов к окислительному стрессу, которому наши клетки подвергаются при старении. В то же время для липидов, участвующих в энергетических процессах, зависимость ровно противоположная. Ненасыщенных энергетических липидов у долгожителей больше, чем у короткоживущих видов. Авторы статьи говорят, что это связано с тем, что такие липиды клеткам сложнее превращать в энергию, а замедленный обмен веществ — одна из характеристик долгоживущих видов.
Екатерина Храмеева, научный сотрудник Сколтеха и ИППИ РАН, один из авторов исследования: «Нам удалось найти связь между продолжительностью жизни вида и особенностями его липидного метаболизма. Этот результат интересен сам по себе, но еще более ценными являются полученные нами данные — липидомы 35 видов млекопитающих. Такой набор данных прекрасно подходит для изучения эволюции липидома, над чем мы и работаем в данный момент. Ранее эволюцию липидома изучали лишь для 4 видов.
Результаты исследования опубликованы в журнале Nature Scientific Reports.
Источник: science.theoryandpractice.ru
Кишечная микрофлора воздействует на наш обмен веществ
23 июня 2018
У системы врожденного иммунитета, нашего первого рубежа защиты от бактериальных инфекций, есть вторая функция, которая также важна, как и первая, и эта функция — точная настройка нашего обмена веществ.
Кишечник фруктовой мушки имеет похожую с человеческим физиологию, только он проще устроен и имеет меньшие размеры.
Ученые говорят, что симбиотические (полезные) бактерии, обитающие в кишечнике, помогают нам в регуляции метаболизма. Новое исследование под руководством Паулы Уотник, доктора медицинских наук из отделения инфекционных заболеваний Бостонской детской больницы, проведенное на плодовых мушках и опубликованное 21 июня 2018 года в «Cell Metabolism» демонстрирует то, как именно наши «хорошие» бактерии это делают.
Пищеварительные клетки кишечника используют врожденные иммунные механизмы для борьбы с вредоносными бактериями. А вот другие клетки кишечника, энтероэндокринные, используют тот же путь (он известен как сигнальный путь IMD) для того, чтобы «хорошие» бактерии реагировали на изменение условий, а именно, подстраивали обмен веществ к диете или к различным видам патологических состояний желудочно-кишечного тракта. «Некоторые врожденные иммунные механизмы существуют не только для иммунитета», — говорит Уотник. «Врожденные иммунные системы кроме этого „слушают“ „хорошие“ бактерии и реагируют на полученную информацию изменением обмена веществ».
Метаболический синдром жировой инфильтрации печени у мух
Уотник и ее коллеги из своего предыдущего исследования знали, что бактерии, живущие в кишечнике мух, создают короткоцепочечную жирную кислоту — ацетат, которая необходима для липидного обмена и для инсулиновой сигнальной системы. Мухи лишенные бактериальной флоры кишечника (следовательно, и ацетата) накапливают жировые капли в пищеварительных клетках.

Определение роли иммунной системы в обмене веществ
Как связаны: кишечная бактерия, врожденная иммунная система и обмен веществ? Через серию экспериментов команда смогла точно выяснить, как именно бактерия оказывает влияние на метаболизм. Они обнаружили следующее:
• Иммунные пути стимулируют энтероэндокринные клетки, чтобы те вырабатывали тахикинин.
• В отсутствие любого из этой пары: бактерии или ее продукта ацетата, тахикинин не производится.
• Когда лишенным кишечной флоры мушкам давали ацетат, врожденные иммунные пути активировались, а обмен веществ нормализовался.
«Мы знаем, что бактерии контролируют наш обмен веществ, но никто не подозревал, что они могут взаимодействовать с врожденными иммунными сигнальными путями в энтероэндокринных клетках», — говорит Уотник.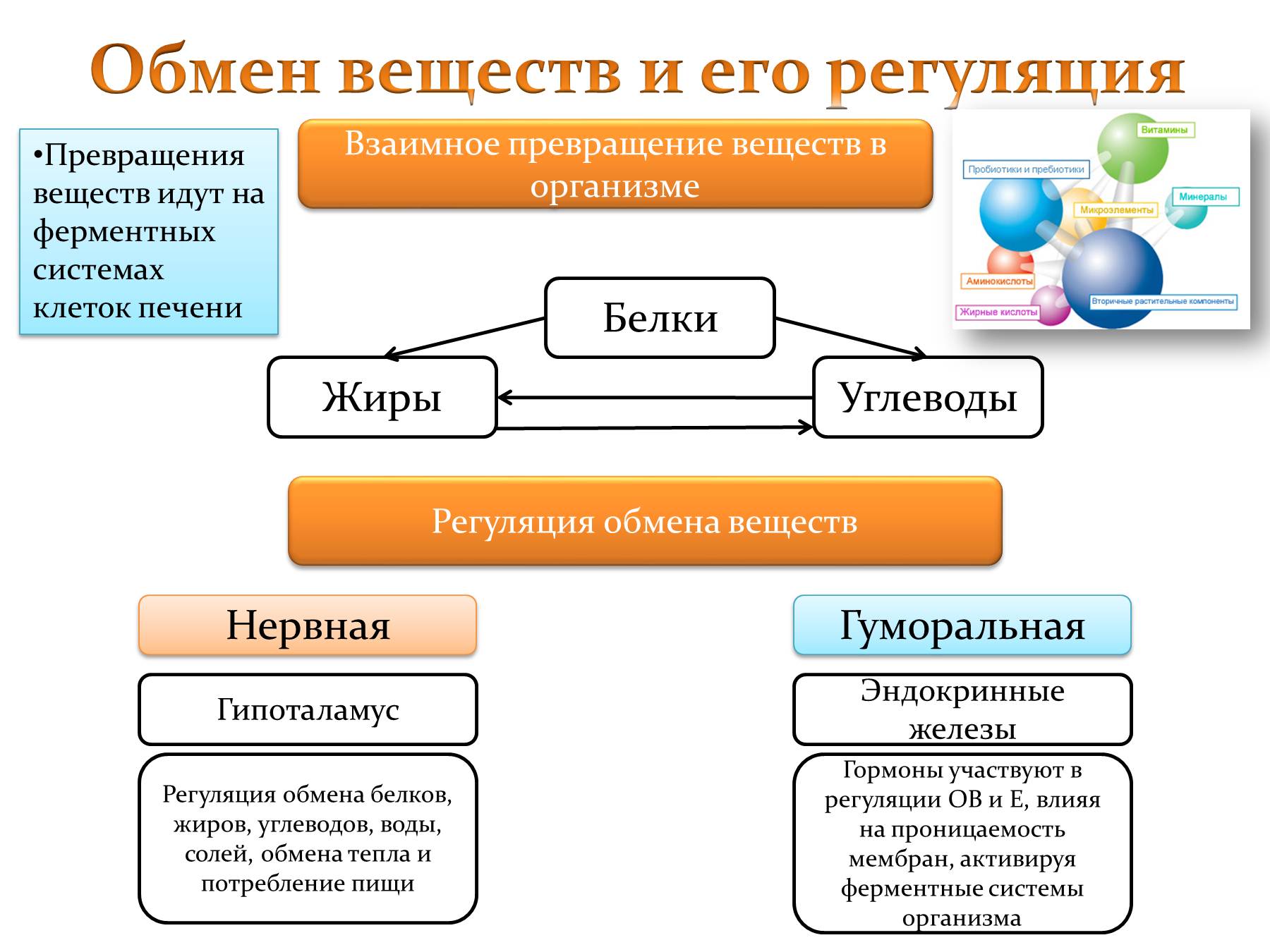 «Возможно, эти пути являются системой, которая позволяет клеткам распознавать бактерии для различных целей».
«Возможно, эти пути являются системой, которая позволяет клеткам распознавать бактерии для различных целей».
Двустороннее взаимодействие
Данная работа также показывает, что активация врожденных иммунных путей в энтероэндокринных клетках является важнейшим фактором для нормального роста и развития мух. Когда Уотник и ее команда инактивировали этот путь, они получали мух с задержкой развития. При скармливании этим особям ацетата или при прямой реактивации у них данного врожденного иммунного пути, их рост возобновлялся.
Если Уотник могла бы сейчас подтвердить полученные результаты на млекопитающих, то дальнейшие исследования обрисовали бы общие черты того, что обеспечивает двустороннее взаимодействие между нашим микробиомом и нашим метаболизмом. «Хорошие» бактерии ферментируют питательные вещества нашей диеты и производят короткоцепочечные жирные кислоты наподобие ацетата, которые помогают нам оптимизировать использование и хранение нутриентов (биологически значимых элементов пищи).
Недосып нарушает обмен веществ — исследование
By Reuters Staff, Рейтер
Сотрудник BMW во время отдыха в специальной комнате, 16 февраля 2011 года. Недосыпание — это не только испорченный день, но и нарушение обмена веществ в человеческом организме, что может привести к набору лишнего веса, показало исследование. REUTERS/Michaela Rehle
НЬЮ-ЙОРК (Рейтер) — Недосыпание — это не только испорченный день, но и нарушение обмена веществ в человеческом организме, что может привести к набору лишнего веса, показало исследование.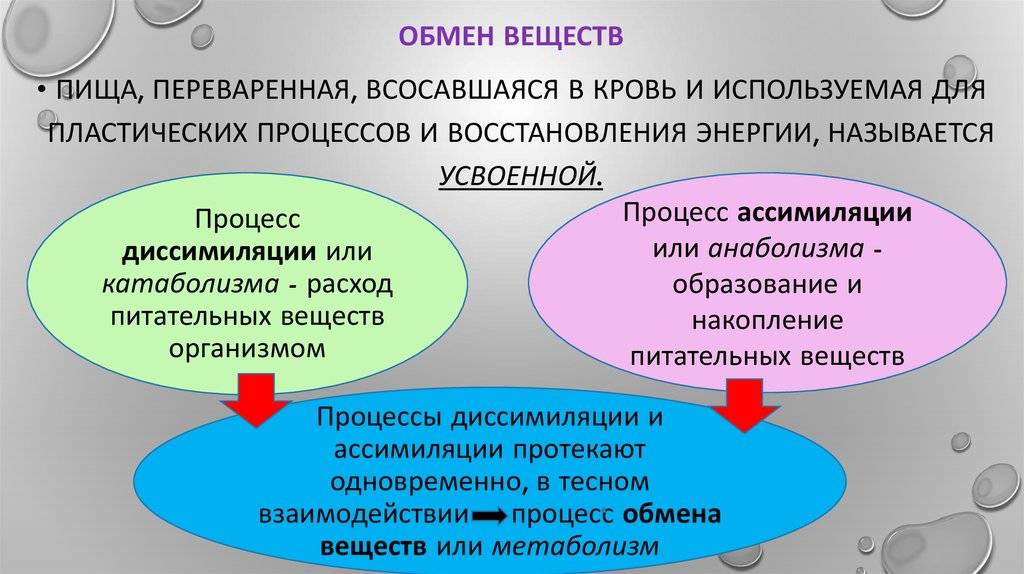
Полученные результаты, опубликованные в American Journal of Clinical Nutrition, стали лишним доказательством того, что недостаток сна может способствовать увеличению веса — не только за счет усиления чувства голода, но также из-за замедления скорости сжигания калорий.
Следовательно увеличение времени сна, по мнению ученых, может предотвратить набор веса.
“Наши изыскания показывают, что одна бессонная ночь резко снижает расход энергии у здоровых мужчин; это свидетельствует, что сон способствует регуляции дневного расхода энергии в организме людей”, — написал руководитель эксперимента Кристиан Бенедикт из Уппсальского университета в Швеции.
Чтобы определить точные механизмы воздействия сна на организм, Бенедикт и его коллеги в течение нескольких дней следили за состоянием 14 студентов мужского пола в разных условиях: при нормальном сне, ограниченном и его полном отсутствии. Затем ученые оценили изменения в количестве потребления пищи, уровне сахара в крови, гормонов и показателей метаболизма.
В ходе исследования обнаружилось, что даже одна ночь, проведенная без сна, замедляет обмен веществ на следующее утро, сокращая расход энергии на выполнение таких задач, как дыхание и пищеварение на 5-20 процентов по сравнению с показателями, полученными наутро после хорошего ночного сна.
При этом уменьшение сна, как выяснилось, не увеличило количество пищи, потребляемой мужчиной в течение дня.
Эксперты из специализированной независимой американской организации National Sleep Foundation рекомендуют взрослым людям спать примерно семь-девять часов каждую ночь.
Наташа Аллен, перевела Яна Соболева
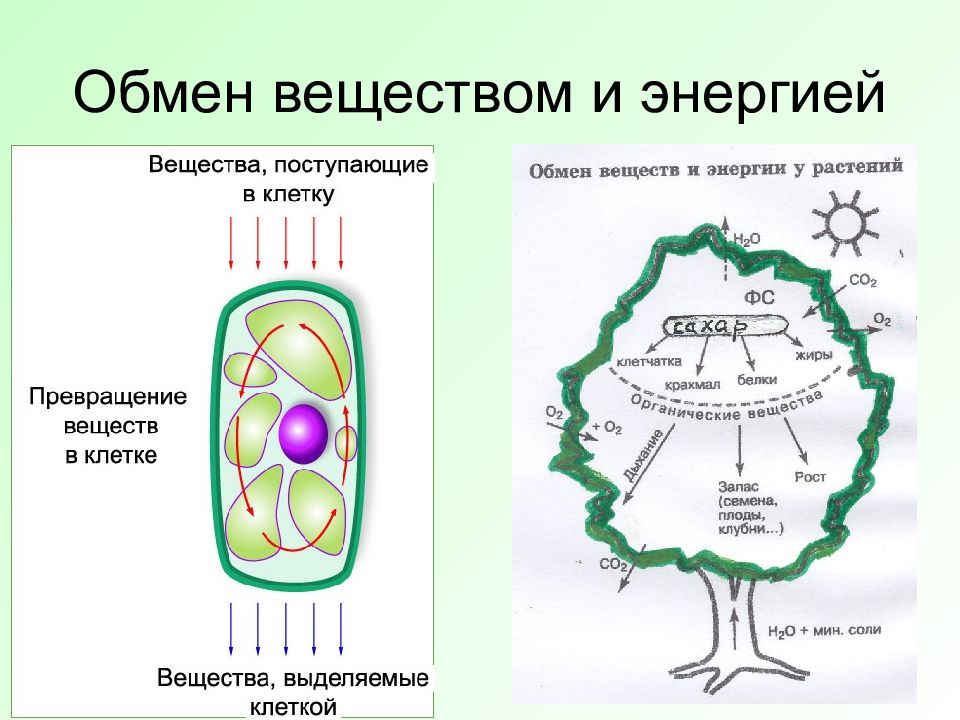
Питание, пищеварение и обмен веществ
Зал посвящён таким важным проблемам физиологии, как питание, пищеварение и обмен веществ. Экспозиция зала даёт возможность узнать и сравнить строение пищеварительной системы человека и животных, знакомит посетителей с особенностями обмена веществ у разных животных, а также позволяет увидеть связь этих процессов с условиями окружающей среды.
В частности, на примере различных видов птиц представлено разнообразие формы клюва в зависимости от типа питания. Ряд экспонатов демонстрирует эволюцию органов захвата пищи (челюстного аппарата) у хордовых животных.
В экспозиции представлен раздел, в котором можно увидеть классический опыт И. П. Павлова. За выдающиеся исследования в области физиологии пищеварения в 1904 году И. П. Павлову была присвоена Нобелевская премия.
Диорама «Шакал и стервятник» показывает важную санитарную роль падальщиков в природе. На диораме «Речные бобры» хорошо видно, что строительная деятельность бобров связана с их питанием. Биогруппа «Мышкующая лиса» демонстрирует сложное поведение хищника, охотящегося за добычей.
Зал посвящён не только пищеварению, но и обмену веществ. Одно из интереснейших явлений в жизни животных — спячка, во время которой у животного падает температура тела, замедляются активные процессы в организме, замедляется обмен веществ. Различают следующие виды спячки: зимний сон, оцепенение, зимнюю и летнюю спячки. В зале представлены биогруппы, демонстрирующие спячку гадюк, бурундука, травяной лягушки, курганчиковой мыши и летучих мышей, что позволяет им переносить неблагоприятные условия среды.
В зале представлены биогруппы, демонстрирующие спячку гадюк, бурундука, травяной лягушки, курганчиковой мыши и летучих мышей, что позволяет им переносить неблагоприятные условия среды.
Небольшой раздел в зале посвящён органам выделения.
Экспозиция зала была открыта в 1988 году. Автор экспозиции — Л. В. Серёгина (Коковкина), художники — Ю. З. Ивантер и В. Я. Грачёв.
В зале проводятся экскурсии «Путь пирожка» для младших школьников и «Пищеварение и обмен веществ» (с демонстрацией опытов) для старшеклассников и студентов.
Генетический тест «Обмен веществ» | Государственное бюджетное учреждение здравоохранения Детская городская клиническая больница (ГБУЗ ДГКБ г. Краснодара)
Пример заключения
«Генетика обмена веществ»
Разработка способов индивидуализации фитнес-программ по контролю веса, основанных на данных генетического анализа является очень популярным направлением. Некоторые из таких первых подходов уже запатентованы и начинают применяться на практике.
Среди генетических факторов, которые могут повлиять на правильный выбор диеты и интенсивности физических нагрузок, чаще всего анализируют нуклеотидные полиморфизмы генов FABP2, PPARG, ADRB2 и ADRB3. Эти гены, а точнее их генетическая вариабельность, значительно влияют на то, как быстро и эффективно усваиваются питательные вещества, поступающие с пищей, а значит, они могут препятствовать, или помогать достижению и поддержке оптимального веса. Эти генетические вариации (полиморфизмы) не являются патогенными мутациями, они достаточно широко распространены, но, располагая информацией об их наличии или отсутствии, можно точнее оценить индивидуальные особенности своего организма.
Так, например, ген FABP2 кодирует белок, связывающий и транспортирующий жирные кислоты в кишечнике. Нуклеотидный вариант («G» или «А») в точке полиморфизма rs1799883 в этом гене приводит к синтезу разных вариантов белка. Один из них лучше связывается с жирными кислотами и, соответственно, люди с таким вариантом гена (генотип GA или AA полиморфизма rs1799883) эффективней усваивают жиры из потребляемой пищи, имеют более высокий индекс массы тела (ИМТ) по сравнению с обладателями альтернативного варианта гена FABP2. Таким пациентам для поддержания оптимального веса рекомендуется питание с пониженным содержанием жиров.
Таким пациентам для поддержания оптимального веса рекомендуется питание с пониженным содержанием жиров.
Ген PPARG, кодирующий гамма-рецептор, активируемый пролифератором пероксисом, отвечает за процессы окисления жирных кислот. Он также влияет на потребность мышечной ткани в глюкозе и ее чувствительность к инсулину. Менее благоприятный вариант полиморфизма rs1801282 (часто называемый как Pro12Ala) приводит к пониженной активности рецептора и, соответственно, к повышению уровня общего холестерина, снижению уровня триглицеридов и повышению чувствительности тканей к инсулину.
Два других популярных гена ADRB2 и ADRB3 кодируют варианты бета-адренергических рецепторов. Встроенные в цитоплазматическую мембрану клетки, они имеют высокую степень сродства к адреналину и регулируют повышение или понижение активности иннервируемой ткани или органа. Активация рецепторов вызывает увеличение интенсивности гликогенолиза в мышцах, увеличение интенсивности секреции инсулина, глюкагона. Полиморфизмы rs1042714 и rs4994 в этих генах влияют на эффективность кодируемых ими рецепторов. При менее благоприятных вариантах легче набирается избыточный вес, а снизить его удается только при более интенсивных тренировках.
Полиморфизмы rs1042714 и rs4994 в этих генах влияют на эффективность кодируемых ими рецепторов. При менее благоприятных вариантах легче набирается избыточный вес, а снизить его удается только при более интенсивных тренировках.
Таким образом, информация по вышеуказанным генетическим полиморфизмам может быть весьма ценной для оптимизации усилий по поддержанию веса и других важных показателей в норме. Варианты генов ADRB2 и ADRB3 могут подсказать то, насколько высокоинтенсивными должны быть физические нагрузки для достижения оптимального результата по коррекции веса. А полиморфизмы генов FABP2 и PPARG позволяют оптимизировать профиль питания.
Тест для определения генетических полиморфизмов
«ОБМЕН ВЕЩЕСТВ»
Результаты теста позволяют подобрать индивидуальную диету при снижении веса, а также определить рекомендуемую интенсивность и тип физических нагрузок.
Питание и обмен веществ у собаки
Что нужно учитывать при выборе рациона для питомца?Обмен веществ у собак имеет ряд особенностей, которые возникли в результате многих веков эволюции их диких предков. Пытаясь что-либо изменить в этих процессах, владельцы неизменно сталкиваются с возникновением разных заболеваний, спровоцированных нарушением обмена веществ.
Пытаясь что-либо изменить в этих процессах, владельцы неизменно сталкиваются с возникновением разных заболеваний, спровоцированных нарушением обмена веществ.
Причиной таких нарушений чаще всего становится неправильно подобранный рацион или несоблюдение правил кормления собаки. Собака по своей природе относится к хищникам, а значит, и состав корма для нее должен быть сбалансирован по белкам, жирам и углеводам в соответствии с ее природой. Так повышенное содержание жиров и углеводов в рационе при отсутствии белка приводит к возникновению нарушения обмена веществ даже у очень молодых собак.
Важно соблюдать режим и периодичность питания. Хищники, к которым относится и собака, начинают охоту, только испытывая сильный голод. А значит, питаются они не три и не два раза в день. Процесс добывания пищи требует огромных энергетических затрат. После удачной охоты хищник наедается и потом долго отдыхает. Так же настроена и система пищеварения домашних собак. Плотная и очень сытная белковая еда, длительный отдых и снова поиск добычи. Поэтому для взрослой здоровой собаки нет надобности дробить суточную норму на очень большое число кормлений. Для щенков и больных собак режим кормления должен быть иным — частота кормления увеличивается при меньшем объеме съедаемого корма.
Поэтому для взрослой здоровой собаки нет надобности дробить суточную норму на очень большое число кормлений. Для щенков и больных собак режим кормления должен быть иным — частота кормления увеличивается при меньшем объеме съедаемого корма.
Еще одна особенность, имеющая важное значение в обмене веществ, — способ поедания пищи. Собака ее не жует и не лакает, а отрывает коренными зубами и заглатывает куски. Крупные куски собака не жует, а также разделяет на более мелкие. Ее желудочно-кишечный тракт приспособлен для переваривания именно такой пищи, поэтому мясо в виде фарша может не усваиваться организмом собаки.
Помимо белка, который собака получает с мясом, для получения дополнительных витаминов и микроэлементов и для лучшего обмена ее организму необходима клетчатка и бактерии. В природе клетчатку и ферменты хищники получают, съев внутренности добычи вместе с содержимым. Для современных собак альтернативой является полностью сбалансированные рационы. Такие рационы незаменимы для собак с нарушениями обмена веществ. Клетчатка не может полностью перевариться организмом собаки, но она меняет фракцию кишечника и стимулирует перистальтику.
Клетчатка не может полностью перевариться организмом собаки, но она меняет фракцию кишечника и стимулирует перистальтику.
Сейчас ветеринарные врачи все чаще сталкиваются с ожирением собак. И дело тут не только в слишком калорийных рационах, содержащих большое количество жиров, а в недостаточно активной жизни домашних собак. Организм хищника привык запасать жир на случай долгого голодания или тяжелой погони за добычей, когда расходуется основная масса его энергетических запасов. Но накопление происходит, а расход не увеличивается, и вскоре владелец сталкивается с нарушением углеводно-жирового баланса организма своей собаки. Поэтому рацион малоподвижной собаки должен быть скорректирован ветеринарным врачом с учетом ее реальной энергозатраты.
Эксперты назвали продукты, ускоряющие метаболизм
Многие люди скептически относятся к утверждению, что после 30 лет худеть гораздо сложнее. Но этому есть вполне логическое научное объяснение – метаболизм становится медленнее, а это напрямую связано с обменом веществ во всем организме. Однако можно помочь своему метаболизму ускориться. Эксперты программы «О самом главном» назвали продукты, которые на самом деле помогут похудеть.
Однако можно помочь своему метаболизму ускориться. Эксперты программы «О самом главном» назвали продукты, которые на самом деле помогут похудеть.
Для того, чтобы расти, человеческий организм должен тратить много энергии. Дети тратят много энергии сами по себе. Примерно в 25 лет все процессы роста в организме начинают замедляться, а после 35 и вовсе останавливаются.
«Мы начинаем катиться под «метаболическую горку», – отметил доктор Сергей Агапкин.
Метаболизм – это процесс обмена веществ в организме. Это та энергия, которая тратится на дыхание, умственную деятельность, поддержание температуры тела, активность. С возрастом скорость метаболизма замедляется примерно на треть. На это влияет состояние поджелудочной железы, мышц, физическая активность и, конечно же, питание. Чтобы ускорить обмен веществ, в свой рацион нужно включить:
Специи — перец и куркуму. Благодаря им повышается общий жизненный тонус и снижается потребность в тяжелой пище и лакомствах.
«Когда проснулся в хорошем настроении, не хочется себя перегружать тяжелой пищей, например, той же колбасой. – объяснил врач-диетолог Михаил Гинзбург. – Вещества, содержащиеся в специях, способствуют тому, что мы больше двигаемся и меньше хотим есть».
Сергей Агапкин добавил, что метаболизм от 10 до 30 процентов от всего обмена – это пищевой термогенез. И он очень чувствителен к острой пище – когда мы едим что-то перченой, наш организм начинает тратить на 10-20 процентов больше калорий.
Ламинария – хороший источник йода, который помогает гормонам ускорять метаболизм. А также. благодаря содержащимся в ней веществах, мы меньше внимания на еду в принципе. Таким образом ламинария не только увеличивает расход энергии, но и нормализует наше питание. Обязательно нужно взять себе на заметку.
Яблочный уксус. Научные исследования показывают, что он повышает чувствительность ткани к инсулину – то есть повышение метаболической пластичности, которая теряется при наборе веса и ожирении. А также уксус снижает гликемический индекс продуктов и помогает снижать вес.
А также уксус снижает гликемический индекс продуктов и помогает снижать вес.
Но существуют и враги метаболизма, о которых не стоит забывать. Сахар, подсолнечное масло, изделия из просеянной муки, картофель и блюда из него (особенно чипсы и пюре), а также колбасные изделия замедляют все процессы в организме. Более того, после такой пищи человеку не хочется двигаться. Не менее важно следить за сном. Чем меньше человек спит, тем больше он начинает есть. Доказано, что таким образом организм восполняет дефицит сна.
Еще больше интересных новостей – в нашем Instagram и Telegram-канале @smotrim_ru.
Центр исследований метаболизма и ожирения
Центр исследований метаболизма и ожирения или CMOR в Институте фундаментальных биомедицинских исследований — это межведомственный и междисциплинарный центр, созданный для поддержки нашего понимания основных биологических механизмов. которые регулируют обмен веществ, и как они нарушаются при таких расстройствах, как ожирение, диабет, инсульт и рак. Хотя может показаться, что это разные темы, у них есть общие коренные причины нарушения энергетического баланса, которые могут повлиять на многие биологические системы.Таким образом, в то время как исследователи CMOR подходят к этим проблемам в рамках своих отдельных дисциплин, Центр предоставляет возможность исследовать общие научные темы и сотрудничать в исследованиях, сочетающих молекулярные и клеточные механизмы метаболизма с широким спектром физиологических и поведенческих исследований. CMOR также работает над тем, чтобы облегчить перевод открытий в прикладные знания для терапии в этих областях.
Хотя может показаться, что это разные темы, у них есть общие коренные причины нарушения энергетического баланса, которые могут повлиять на многие биологические системы.Таким образом, в то время как исследователи CMOR подходят к этим проблемам в рамках своих отдельных дисциплин, Центр предоставляет возможность исследовать общие научные темы и сотрудничать в исследованиях, сочетающих молекулярные и клеточные механизмы метаболизма с широким спектром физиологических и поведенческих исследований. CMOR также работает над тем, чтобы облегчить перевод открытий в прикладные знания для терапии в этих областях.
Начиная с исследований митохондриального метаболизма Альберта Ленингера и заканчивая работами Питера Педерсена в области биоэнергетики и многими другими, Университет Джона Хопкинса создал выдающееся наследие открытий в области метаболизма и системной биологии.Рассматривая такие темы, как определение питательных веществ, биоэнергетика и эндокринная регуляция, центр использует как передовые технологии, так и фундаментальную фундаментальную науку, чтобы углубить наше понимание биологии, которая регулирует метаболизм, и того, как он нарушается при сопутствующих расстройствах, таких как ожирение, диабет, рак, и инсульт.
Директор: Михаэль Вольфганг, доктор философии.
Миссия
Возглавлять и поддерживать комплексные исследования в области метаболизма и ожирения, чтобы углубить наше понимание биологических механизмов, регулирующих обмен веществ, и того, как они нарушаются при сопутствующих расстройствах, таких как ожирение и диабет.
Видение: Понимание того, как определенные метаболические пути влияют на биологические результаты и поведение, является целью метаболизма в нашу эпоху и является общей основой для систем и поведенческой биологии. Общая цель CMOR состоит в том, чтобы разработать инфраструктуру для проведения передовых исследований в области фундаментальной фундаментальной науки о метаболизме.
Цели
- Обеспечить инфраструктуру для научного взаимодействия между преподавателями и сообществом.
- Для интеграции исследований с использованием модельных организмов и метаболического профиля.
- Развитие сервисных и технологических ресурсов.

- Для повышения уровня образования стажеров.
- Содействовать взаимодействию между CMOR и агентствами, которые поддерживают исследования в области метаболизма и ожирения.
- Развитие сервисных и технологических ресурсов.
- Распространение знаний среди населения в форме последипломного образования и содействие использованию этих знаний в терапевтических стратегиях.
- Программные темы:
- Регуляция/восприятие питательных веществ охватывает метаболические пути, связанные с распределением углеводов, глюкозы и липидов, которые влияют на то, как энергия используется для поддержки синтеза макромолекул.
- Биоэнергетика включает преобразование катаболизированных молекул в энергию (АТФ) и восстановительные эквиваленты (НАДН), доступность которых диктует возможные биологические анаболические процессы.
- Эндокринная регуляция означает гуморальные и нейроэндокринные реакции, которые происходят для поддержания гомеостаза, часто в связи с доступностью питательных веществ. Например, такие молекулы, как инсулин, лептин и гипоталамические нейропептиды, реагируют на периферические и центральные метаболические сигналы, чтобы ощущать энергетический баланс и влиять на поведение. На клеточном уровне эти метаболические системы влияют на выживаемость клеток, регуляцию клеточного цикла, уровни экспрессии различных белков, необходимых для клеточных функций, и клеточное старение. На уровне организма питательные вещества, эндокринные профили и биоэнергетика влияют на воспроизводство, переносимость физических нагрузок, активность ЦНС, пищевое поведение и продолжительность жизни.
- Программные темы:

 На клеточном уровне эти метаболические системы влияют на выживаемость клеток, регуляцию клеточного цикла, уровни экспрессии различных белков, необходимых для клеточных функций, и клеточное старение. На уровне организма питательные вещества, эндокринные профили и биоэнергетика влияют на воспроизводство, переносимость физических нагрузок, активность ЦНС, пищевое поведение и продолжительность жизни.
На клеточном уровне эти метаболические системы влияют на выживаемость клеток, регуляцию клеточного цикла, уровни экспрессии различных белков, необходимых для клеточных функций, и клеточное старение. На уровне организма питательные вещества, эндокринные профили и биоэнергетика влияют на воспроизводство, переносимость физических нагрузок, активность ЦНС, пищевое поведение и продолжительность жизни.Нарушение регуляции этих путей приводит к некоторым из самых разрушительных заболеваний, с которыми мы сталкиваемся, включая ожирение, диабет, рак и инсульт.По мере того, как мы обнаруживаем новые регулирующие роли для этих метаболических путей, мы стремимся применить эту информацию к терапевтическим стратегиям, что делает Центр своевременным мероприятием. Самое главное, эта тема обеспечивает гибкость, позволяя Центру при необходимости адаптироваться к будущим потребностям.
Основные услуги в области физиологии животных
CMOR создала основной сервисный центр Coordinated Services for Metabolism Research (CSMR) для выполнения своей миссии по содействию совместным исследованиям в области метаболизма. С 2008 года CSMR предоставляет доступ к общим инструментам, тестовому пространству и опыту для метаболического фенотипирования in vivo на моделях грызунов сообществу Университета Джона Хопкинса и за его пределами. Сьюзен Аджа, доктор философии, руководит CSMR, курирует и проводит исследования, предоставляет экспертные знания и обучение, а также направляет исследователей к дополнительным ресурсам для поддержки их проектов.
С 2008 года CSMR предоставляет доступ к общим инструментам, тестовому пространству и опыту для метаболического фенотипирования in vivo на моделях грызунов сообществу Университета Джона Хопкинса и за его пределами. Сьюзен Аджа, доктор философии, руководит CSMR, курирует и проводит исследования, предоставляет экспертные знания и обучение, а также направляет исследователей к дополнительным ресурсам для поддержки их проектов.
Факультет
Исследовательский факультет
- Лили Айяла Баруш, доктор медицины
- Тодд Тарквин Браун, М.Д., к.т.н.
- Адриан Сандра Добс, M.D., M.H.S.
- Питер Эспеншейд, доктор философии.
- Даниэль М. Рабен, доктор философии.
- Дипали Шарма, MS, Ph.D.
- Михаэль Вольфганг, доктор философии.
- Гуанг Уильям Вонг, доктор философии.
- Джеффри Хантер Янг, M.D., M.H.S.
- Хэн Чжу, доктор философии.
Дополнительный факультет
Сьюзан Аджа
Департамент неврологии
Сьюзан Аджа привносит большой опыт в исследования, связанные с метаболизмом, и управление проектами в качестве директора CMOR. Исследования (CSMR).Она получила степень бакалавра и доктора физиологии в Калифорнийском университете в Дэвисе, провела постдокторскую подготовку в области поведенческой неврологии в Медицинской школе Университета Джона Хопкинса, а с 2002 года преподает на медицинском факультете. нейротрансмиттеры периферической нервной системы, нейропептиды, ампер-активируемая протеинкиназа, модуляция метаболизма жирных кислот и цикл ТСА/здоровье митохондрий в регуляции энергетического баланса при метаболическом здоровье и заболевании.Дополнительная деятельность включает в себя ее роль с 2015 года в качестве связного с клиентами для ядра молекулярных детерминантов в детской больнице Джона Хопкинса. Доктор Айя стремится направлять исследователей к оптимальным ресурсам и опыту, а также координировать проекты для достижения успешных результатов.
Исследования (CSMR).Она получила степень бакалавра и доктора физиологии в Калифорнийском университете в Дэвисе, провела постдокторскую подготовку в области поведенческой неврологии в Медицинской школе Университета Джона Хопкинса, а с 2002 года преподает на медицинском факультете. нейротрансмиттеры периферической нервной системы, нейропептиды, ампер-активируемая протеинкиназа, модуляция метаболизма жирных кислот и цикл ТСА/здоровье митохондрий в регуляции энергетического баланса при метаболическом здоровье и заболевании.Дополнительная деятельность включает в себя ее роль с 2015 года в качестве связного с клиентами для ядра молекулярных детерминантов в детской больнице Джона Хопкинса. Доктор Айя стремится направлять исследователей к оптимальным ресурсам и опыту, а также координировать проекты для достижения успешных результатов.
Библиография: https://pubmed.ncbi.nlm.nih.gov/?term=Aja+S&cauthor_id=32313611
Контактная информация:
855 Н. Вулф Стрит
Балтимор 473 9000 9000
Тел. : 443-287-7671
: 443-287-7671
Факс: 410-614-8033
Мишель Шермак
Отделение хирургии
, Михеле ШермакД., заведующий отделением пластической хирургии в Медицинском центре Джонса Хопкинса Бэйвью, а также член Центра бариатрической хирургии. Клинически она специализируется на новейших методах эстетической хирургии и хирургии груди. Ее исследования сосредоточены на фундаментальной науке о заживлении мягких тканей, заживлении костей и тканевой инженерии, а также она интересуется влиянием потери веса на физиологию и метаболизм адипоцитов.
Эл.
Обновлено 28 октября 2021 г.
Правда в том, что погоня за быстрым метаболизмом может не помочь с потерей веса.Но вы можете улучшить свое здоровье и немного улучшить свой метаболизм с помощью этих простых методов.
Представьте себе легкость похудения, если бы это было так же просто, как щелкнуть выключателем — и вот так ваше тело волшебным образом сжигает энергию более эффективно. В рекламе 1 в Instagram и на полках продуктовых магазинов представлено множество продуктов, которые обещают сделать именно это, повысив метаболизм вашего тела. Но реальность такова, что метаболизм намного сложнее.
В рекламе 1 в Instagram и на полках продуктовых магазинов представлено множество продуктов, которые обещают сделать именно это, повысив метаболизм вашего тела. Но реальность такова, что метаболизм намного сложнее.
Как работает метаболизм?
Метаболизм каждого человека — процесс, посредством которого организм превращает пищу в энергию 2 — во многом определяется их генами.Таким образом, ваш метаболизм является более или менее заданным значением, и у разных людей он будет разным. 3
«Даже когда мы вообще ничего не делаем, обмен веществ все равно будет поддерживать функционирование организма на так называемой базовой скорости метаболизма. Этот показатель варьируется от человека к человеку», — говорит Эмили Вундер, MSCN, RD, LDN, зарегистрированный диетолог и создатель Healthier Taste, веб-сайта питательных рецептов. «Итак, хотя это может показаться несправедливым, человек рядом с вами может естественным образом сжигать больше калорий, чем вы, в то время как вы оба ничего не делаете.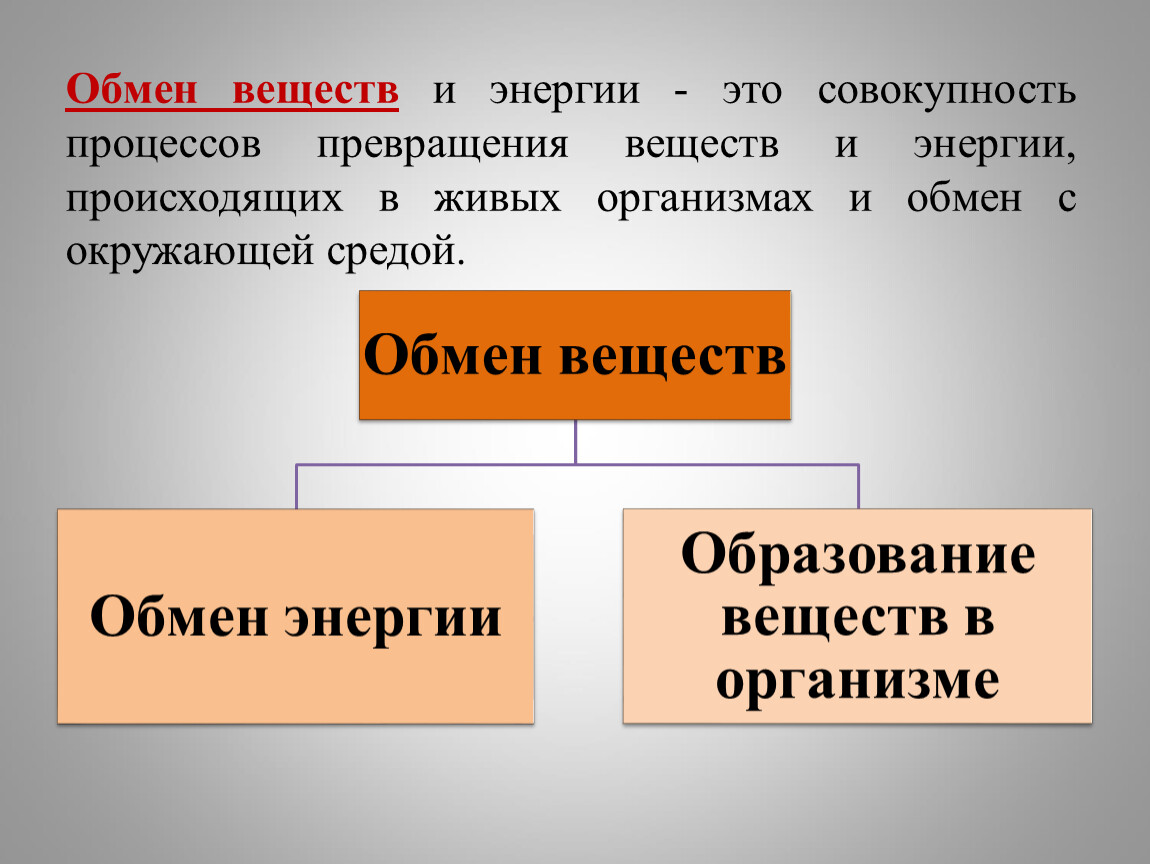 К другим факторам, влияющим на обмен веществ, относятся старение (обмен веществ замедляется с возрастом) и пол, поскольку у мужчин, как правило, скорость метаболизма выше, чем у женщин. 4
К другим факторам, влияющим на обмен веществ, относятся старение (обмен веществ замедляется с возрастом) и пол, поскольку у мужчин, как правило, скорость метаболизма выше, чем у женщин. 4
Потеря веса и обмен веществ
Ирония в том, что метаболизм можно рассматривать как средство для снижения веса, поскольку усилия по снижению веса могут на самом деле замедлить ваш метаболизм. «Один из очень распространенных способов, которым это происходит, — это когда люди пропускают приемы пищи или отказываются от еды в течение длительного периода времени, чтобы похудеть», — говорит Вундер. Сначала вы можете похудеть, но когда ваше тело будет лишено еды, оно начнет реагировать соответствующим образом.«Наши тела умны, — говорит Вундер. «Когда мы находимся в состоянии голодания, тело будет удерживать жир, а не сжигать его, и со временем это приводит к накоплению большего количества жира и замедлению скорости метаболизма».
Как добиться здорового обмена веществ
Вместо того, чтобы думать о метаболизме как о быстром средстве для похудения (подсказка: быстрых решений на самом деле не существует), попробуйте вместо этого сосредоточиться на своем здоровье. Изменения в вашем образе жизни могут помочь немного увеличить ваш метаболизм, но, что более важно, улучшит ваше самочувствие и поможет вам стать лучше, а это более важно, чем потеря веса.Ведь ты не цифра на весах. Ваш размер брюк? Это действительно не имеет значения. Попробуйте эти техники, чтобы почувствовать себя более энергичным, отдохнувшим и здоровым, независимо от того, быстрый у вас метаболизм или нет.
Изменения в вашем образе жизни могут помочь немного увеличить ваш метаболизм, но, что более важно, улучшит ваше самочувствие и поможет вам стать лучше, а это более важно, чем потеря веса.Ведь ты не цифра на весах. Ваш размер брюк? Это действительно не имеет значения. Попробуйте эти техники, чтобы почувствовать себя более энергичным, отдохнувшим и здоровым, независимо от того, быстрый у вас метаболизм или нет.
Упражнения и обмен веществ. Посещение тренажерного зала или занятия спортом дома — отличный способ оставаться в форме и оставаться здоровым. Но каждое небольшое движение помогает, особенно в нашей преимущественно сидячей культуре. «Помимо преднамеренных упражнений, общий уровень активности может играть роль в вашем метаболизме», — говорит Вундер.«Если вы будете больше двигаться в течение дня небольшими способами, со временем эти действия будут складываться и поддерживать метаболизм». Попробуйте парковаться далеко от двери продуктового магазина, подниматься по лестнице, ходить пешком во время обеденного перерыва и делать растяжку в течение дня. 5
5
Тяжелая атлетика и обмен веществ. Исследования показали, что силовые тренировки помогают сохранить мышечную массу. «Более высокая мышечная масса связана с более высокой скоростью метаболизма», — объясняет Вундер. Поскольку мышечная масса уменьшается с возрастом, то же самое может происходить и с обменом веществ, не говоря уже о силе.Так что попробуйте включить поднятие тяжестей в свою тренировочную программу, чтобы оценить все ее преимущества. 6
Сон и обмен веществ. Ни для кого не секрет, что достаточное количество сна ночью является ключом к сохранению здоровья, и это также касается вашего метаболизма. Качественный сон поможет поддерживать скорость метаболизма. Одно исследование показало, что всего несколько ночей недосыпа могут заставить людей чувствовать себя менее сытыми после еды. Это также показало, что люди, лишенные сна, по-разному усваивают жир в пище. 7 Так что отдыхай! Ваше тело скажет вам спасибо.
Усилители метаболизма. Наконец, помните, что продукты, предназначенные для ускорения обмена веществ, не являются чудодейственным средством. «Ажиотаж вокруг этих продуктов кажется большим, но исследования, связанные с ними, ограничены», — говорит Вундер. «Многие из этих продуктов содержат вещества, которые уже есть в нашем организме, такие как аминокислоты и минералы. Добавление большего их количества не дает убедительных результатов в отношении влияния на метаболизм и вес». Кофеин является распространенным ингредиентом, который может влиять на обмен веществ, но некоторые уровни кофеина в таких продуктах могут быть небезопасными.Wunder рекомендует проконсультироваться с врачом, прежде чем пробовать такие продукты.
В конечном счете, лучшее, что вы можете сделать для себя, — это перестать беспокоиться о своем метаболизме и просто вести здоровый образ жизни. «Глупо напрягаться из-за того, что на самом деле нельзя изменить», — говорит Вундер. «Не существует фиксированного переключателя для ускорения метаболизма, но изменение образа жизни с течением времени оказывает влияние».
Кредитная карта CareCredit — это простой способ оплаты медицинских и оздоровительных услуг, не покрываемых страховкой.Воспользуйтесь локатором принятия или загрузите мобильное приложение CareCredit, чтобы найти поставщика или продавца рядом с вами, который принимает CareCredit.
Содержание может быть изменено без предварительного уведомления и предоставляется исключительно для вашего удобства. Вам настоятельно рекомендуется проконсультироваться с вашими индивидуальными консультантами и/или поставщиками медицинских услуг в отношении любой представленной информации. Ни Synchrony, ни какие-либо из ее аффилированных лиц, включая CareCredit, не делают никаких заявлений или гарантий в отношении этого контента и не несут ответственности за любые убытки или ущерб, возникшие в результате использования предоставленной информации.
Адаптации в метаболизме и трансляции белков приводят к эффекту Крэбтри у дрожжей
Значение
Избыточный метаболизм, известный как эффект Крэбтри у дрожжей, представляет собой кажущуюся расточительной стратегию использования аэробного брожения вместо более эффективного дыхания для выработки энергии . Это позволяет клеткам расти быстрее при высокой доступности глюкозы. Несмотря на то, что они хорошо изучены, основные причины того, что не все дрожжи испытывают эффект Крэбтри, но при этом способны расти со скоростью, сравнимой с дрожжами, проявляющими этот эффект, неизвестны.Мы объединили количественную оценку физиологии и протеома с метаболическим моделированием, чтобы провести количественное сравнение четырех дрожжей, двух проявляющих и двух не проявляющих эффект Крэбтри, в условиях избытка глюкозы. Наш анализ дает представление об основных причинах эффекта Крэбтри, демонстрируя связь с адаптацией как в метаболизме, так и в трансляции белков.
Это позволяет клеткам расти быстрее при высокой доступности глюкозы. Несмотря на то, что они хорошо изучены, основные причины того, что не все дрожжи испытывают эффект Крэбтри, но при этом способны расти со скоростью, сравнимой с дрожжами, проявляющими этот эффект, неизвестны.Мы объединили количественную оценку физиологии и протеома с метаболическим моделированием, чтобы провести количественное сравнение четырех дрожжей, двух проявляющих и двух не проявляющих эффект Крэбтри, в условиях избытка глюкозы. Наш анализ дает представление об основных причинах эффекта Крэбтри, демонстрируя связь с адаптацией как в метаболизме, так и в трансляции белков.
Abstract
Аэробное брожение, также называемое эффектом Крэбтри у дрожжей, является хорошо изученным явлением, которое позволяет многим эукариальным клеткам достигать более высоких скоростей роста при высокой доступности глюкозы.Не все дрожжи проявляют эффект Крэбтри, и неизвестно, почему Крэбтри-отрицательные дрожжи могут расти со скоростью, сравнимой с Крэбтри-положительными дрожжами. Здесь мы количественно сравнили два Крэбтри-положительных дрожжей, Saccharomyces cerevisiae и Schizosaccharomyces pombe , и два Крэбтри-отрицательных дрожжей, Kluyveromyces marxianus и Schefferomyces stipitis , культивируемых. Объединив количественную оценку физиологии и протеома с метаболическим моделированием в масштабе генома, мы обнаружили, что две группы различаются по энергетическому метаболизму и эффективности трансляции.У Crabtree-позитивных дрожжей поток центрального углеродного метаболизма и распределение протеома благоприятствуют стратегии утилизации глюкозы, сводящей к минимуму стоимость протеома, поскольку параметры трансляции белков, включая рибосомное содержание и/или эффективность, ниже. Crabtree-негативные дрожжи, однако, используют стратегию максимизации выхода АТФ, что сопровождается более высокими параметрами трансляции белка. Наш анализ дает представление об основных причинах эффекта Крэбтри, демонстрируя связь с адаптацией как в метаболизме, так и в трансляции белков.
Здесь мы количественно сравнили два Крэбтри-положительных дрожжей, Saccharomyces cerevisiae и Schizosaccharomyces pombe , и два Крэбтри-отрицательных дрожжей, Kluyveromyces marxianus и Schefferomyces stipitis , культивируемых. Объединив количественную оценку физиологии и протеома с метаболическим моделированием в масштабе генома, мы обнаружили, что две группы различаются по энергетическому метаболизму и эффективности трансляции.У Crabtree-позитивных дрожжей поток центрального углеродного метаболизма и распределение протеома благоприятствуют стратегии утилизации глюкозы, сводящей к минимуму стоимость протеома, поскольку параметры трансляции белков, включая рибосомное содержание и/или эффективность, ниже. Crabtree-негативные дрожжи, однако, используют стратегию максимизации выхода АТФ, что сопровождается более высокими параметрами трансляции белка. Наш анализ дает представление об основных причинах эффекта Крэбтри, демонстрируя связь с адаптацией как в метаболизме, так и в трансляции белков.
Рост является фундаментальным свойством жизни, поэтому основной задачей организмов является поглощение и расщепление питательных веществ из окружающей среды для выработки энергии в форме аденозинтрифосфата (АТФ) и синтеза строительных блоков, необходимых для размножения. Многие организмы используют глюкозу в качестве предпочтительного источника углерода и энергии. Существует две основные стратегии метаболизма глюкозы: 1) полное окисление глюкозы до углекислого газа и воды с использованием потребляющего кислород дыхательного пути, состоящего из гликолиза, цикла трикарбоновых кислот (TCA) и окислительного фосфорилирования, и 2) неполное окисление посредством гликолиза и ферментация с выделением побочных продуктов.С точки зрения энергии, вырабатываемой на молекулу глюкозы, полное окисление глюкозы посредством дыхания примерно в 10 раз эффективнее, чем неполное окисление посредством ферментации, а выход биомассы в пять раз выше (1⇓–3). Несмотря на это, использование ферментативных путей для метаболизма глюкозы с высокой скоростью роста наблюдается у многих организмов, от бактерий и дрожжей до клеток человека (4⇓–6). Это явление называется избыточным метаболизмом и характеризуется высокими потоками гликолиза, повышенным направлением углерода в сторону побочных продуктов брожения и снижением потока через дыхание даже в присутствии кислорода.
Это явление называется избыточным метаболизмом и характеризуется высокими потоками гликолиза, повышенным направлением углерода в сторону побочных продуктов брожения и снижением потока через дыхание даже в присутствии кислорода.
Существует множество теорий основных причин избыточного метаболизма (7). Пфайффер и соавт. (3) предположили, что переполнение является результатом компромисса между скоростью и выходом синтеза АТФ двух метаболических режимов. Недавние исследования расширили эту идею, включив в нее стоимость белка, предполагая, что избыточный метаболизм вызван ограниченными ресурсами протеома. Поскольку содержание белка в клетке ограничено, существует естественное ограничение на долю протеома, которая может быть выделена для метаболизма.Хотя дыхание имеет высокий выход АТФ, оно требует синтеза компонентов цикла ТСА и цепи переноса электронов (ЭТЦ), а также дополнительного митохондриального механизма для эукариальных клеток, в то время как для ферментации требуется лишь несколько цитозольных ферментов. Следовательно, при высоких скоростях роста ферментация более эффективна с точки зрения массы протеома, необходимой для производства того же количества АТФ. Действительно, различные подходы к моделированию (1, 7⇓–9) и протеомные измерения (10) показали, что, когда позволяют условия, например, когда глюкоза присутствует в изобилии, клетки будут участвовать в избыточном метаболизме, позволяя им расти с большей скоростью. .Для микроорганизмов, таких как бактерии и дрожжи, это дает эволюционное преимущество, позволяющее клеткам превзойти своих соседей.
Следовательно, при высоких скоростях роста ферментация более эффективна с точки зрения массы протеома, необходимой для производства того же количества АТФ. Действительно, различные подходы к моделированию (1, 7⇓–9) и протеомные измерения (10) показали, что, когда позволяют условия, например, когда глюкоза присутствует в изобилии, клетки будут участвовать в избыточном метаболизме, позволяя им расти с большей скоростью. .Для микроорганизмов, таких как бактерии и дрожжи, это дает эволюционное преимущество, позволяющее клеткам превзойти своих соседей.
У дрожжей избыточный метаболизм называют эффектом Крэбтри (11, 12), но интересно, что не все виды дрожжей проявляют избыточный метаболизм. Некоторые дрожжи могут полностью окислять глюкозу за счет дыхания даже при высоких концентрациях глюкозы (Crabtree-негативные дрожжи) и при этом расти со скоростью, сравнимой со скоростью Crabtree-позитивных дрожжей.Механизм этого не ясен. Поэтому мы провели абсолютную количественную оценку протеома двух Crabtree-положительных дрожжей, Saccharomyces cerevisiae и Schizosaccharomyces pombe , и двух Crabtree-негативных дрожжей, Kluyveromyces marxianus и Schefferomyces stipitis . Мы объединили количественную оценку физиологии и протеома с метаболическим моделированием в масштабе генома (GEM), чтобы количественно описать различия между Crabtree-позитивными и -негативными дрожжами как на протеомном, так и на метаболическом уровнях.Мы показываем, что эффект Крэбтри связан с различиями метаболических потоков в гликолизе, пентозофосфатном пути (ПФП) и дыхательных потоках. Далее мы обнаружили, что распределение протеома отражает компромисс между двумя различными стратегиями утилизации глюкозы у Crabtree-позитивных и -негативных дрожжей, с ограничениями метаболической способности, особенно в ETC и АТФ-синтазе. Наконец, мы показываем, что эффект Крэбтри сопровождается различиями в содержании рибосомного белка (РП) и эффективности трансляции белка.Взятые вместе, наши анализы показывают, что в основе эффекта Крэбтри у дрожжей лежит адаптация как метаболизма, так и трансляции белков.
Мы объединили количественную оценку физиологии и протеома с метаболическим моделированием в масштабе генома (GEM), чтобы количественно описать различия между Crabtree-позитивными и -негативными дрожжами как на протеомном, так и на метаболическом уровнях.Мы показываем, что эффект Крэбтри связан с различиями метаболических потоков в гликолизе, пентозофосфатном пути (ПФП) и дыхательных потоках. Далее мы обнаружили, что распределение протеома отражает компромисс между двумя различными стратегиями утилизации глюкозы у Crabtree-позитивных и -негативных дрожжей, с ограничениями метаболической способности, особенно в ETC и АТФ-синтазе. Наконец, мы показываем, что эффект Крэбтри сопровождается различиями в содержании рибосомного белка (РП) и эффективности трансляции белка.Взятые вместе, наши анализы показывают, что в основе эффекта Крэбтри у дрожжей лежит адаптация как метаболизма, так и трансляции белков.
Результаты
Характеристика физиологических различий между Крэбтри-положительными и -отрицательными дрожжами.

Мы выбрали S. cerevisiae и S. pombe в качестве репрезентативных Крэбтри-положительных дрожжей, поскольку они широко используются в качестве модельных эукариалов и независимо развили эффект Крэбтри (13). В качестве Crabtree-негативных дрожжей мы выбрали K.marxianus и S. stipitis , которые оказались привлекательными для использования в качестве клеточных фабрик (14, 15). Чтобы охарактеризовать физиологию эффекта Крэбтри, мы культивировали все четыре вида дрожжей в трех повторностях в биореакторах в минимальной среде (16) с глюкозой в качестве источника углерода и измеряли важные физиологические параметры, включая биомассу, уровни экзометаболитов, а также поглощение кислорода и содержание углерода. производство двуокиси (рис. 1 A – D ). С момента переполнения метаболизма в К.marxianus и S. stipitis возникает в ответ на ограничение доступности кислорода (14, 17), мы контролировали уровни растворенного кислорода (DO), чтобы они всегда были выше 60% ( Приложение SI , рис. S1). ), а начальная концентрация глюкозы у K. marxianus составляла 10 г/л вместо 20 г/л, как для других видов. Как видно из профиля экзометаболитов, у двух Crabtree-негативных дрожжей наблюдалось низкое образование побочных продуктов (рис. 1 A , B и E и SI Приложение , рис.S2 A и B ). K. marxianus выделял небольшое количество ацетата, что соответствует <3% потребляемой глюкозы в соответствии с предыдущими исследованиями K. marxianus во время периодического культивирования (17, 18). Крэбтри-положительные виды секретировали более высокие уровни побочных продуктов с этанолом в качестве основного продукта, что соответствует 42 и 47% потребления глюкозы для S. pombe и S. cerevisiae соответственно (рис. 1 C). – Е ).Мы также наблюдали большую разницу в скорости поглощения глюкозы (GUR) между группами (рис. 1 F и SI Приложение , таблица S1).
S1). ), а начальная концентрация глюкозы у K. marxianus составляла 10 г/л вместо 20 г/л, как для других видов. Как видно из профиля экзометаболитов, у двух Crabtree-негативных дрожжей наблюдалось низкое образование побочных продуктов (рис. 1 A , B и E и SI Приложение , рис.S2 A и B ). K. marxianus выделял небольшое количество ацетата, что соответствует <3% потребляемой глюкозы в соответствии с предыдущими исследованиями K. marxianus во время периодического культивирования (17, 18). Крэбтри-положительные виды секретировали более высокие уровни побочных продуктов с этанолом в качестве основного продукта, что соответствует 42 и 47% потребления глюкозы для S. pombe и S. cerevisiae соответственно (рис. 1 C). – Е ).Мы также наблюдали большую разницу в скорости поглощения глюкозы (GUR) между группами (рис. 1 F и SI Приложение , таблица S1). S. pombe и S. cerevisiae имели примерно в два и более раза более высокую скорость поглощения, чем виды, отрицательные по Крэбтри, соответственно. Это согласуется с наблюдением, что GUR играет важную роль в избыточном метаболизме (19).
S. pombe и S. cerevisiae имели примерно в два и более раза более высокую скорость поглощения, чем виды, отрицательные по Крэбтри, соответственно. Это согласуется с наблюдением, что GUR играет важную роль в избыточном метаболизме (19).
Характеристика физиологических различий между Crabtree-позитивными и -негативными дрожжами.( A – D ) Плотность клеток (оптическая плотность при 600 нм [OD600]) и внеклеточные концентрации глюкозы, этанола, глицерина и ацетата в ходе периодического культивирования. Показаны средние значения ± стандартное отклонение биологических троек. Дополнительные метаболиты см. в приложении SI , рис. S2 A – D . ( E ) Фракционное потребление глюкозы, С-ммоль/С-моль глюкозы, нормализованное по отношению к общему извлечению углерода. Показаны средние значения биологических троек.Извлечение углерода составляло от 91 до 100% для всех четырех дрожжей. ( Ф ) ГУР. Показаны средние значения ± стандартное отклонение биологических троек. ( G ) Содержание белка (P tot ), выход биомассы (Y sx ) и скорость роста (µ) в зависимости от GUR. Показаны средние значения ± стандартное отклонение и индивидуальные значения биологических троек. Цветные прямоугольники представляют организм, как указано на панели F . ( H ) График OUR, CER и RQ в зависимости от GUR. Средние значения ± стандартное отклонение и индивидуальные значения биологических троек показаны для S.stipitis и средние значения, а также индивидуальные значения для биологических дубликатов показаны для S. pombe , S. cerevisiae и K. marxianus . Цветные прямоугольники представляют организм, как показано на панели F . Spo, S. pombe ; Sce, S. cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis.
( G ) Содержание белка (P tot ), выход биомассы (Y sx ) и скорость роста (µ) в зависимости от GUR. Показаны средние значения ± стандартное отклонение и индивидуальные значения биологических троек. Цветные прямоугольники представляют организм, как указано на панели F . ( H ) График OUR, CER и RQ в зависимости от GUR. Средние значения ± стандартное отклонение и индивидуальные значения биологических троек показаны для S.stipitis и средние значения, а также индивидуальные значения для биологических дубликатов показаны для S. pombe , S. cerevisiae и K. marxianus . Цветные прямоугольники представляют организм, как показано на панели F . Spo, S. pombe ; Sce, S. cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis.
Основные параметры роста представлены на рис. 1 G и H . Измеренная скорость роста была аналогичной для S. cerevisiae и двух Crabtree-негативных видов, 0,42 ч -1 по сравнению с 0,44 и 0,47 ч -1 для K. marxianus и S. stipitis соответственно, в то время как при S. pombe ч -1 . Ферментативный метаболизм у Crabtree-положительных дрожжей отражался в выходе биомассы (Y sx ) около 0,1 г сухого веса (г DW)/г глюкозы и дыхательных коэффициентах (RQ) около 9. Дыхательный метаболизм Crabtree-отрицательных дрожжей вид был подтвержден значениями RQ 1.09 и 1,15 для K. marxianus и S. stipitis соответственно, что привело к выходу биомассы 0,44 и 0,58 г DW/г глюкозы соответственно. Интересно, что мы наблюдали различия в скорости поглощения кислорода (OUR) и скорости выделения углекислого газа (CER) при сравнении видов в каждой группе. Для Crabtree-негативных видов S. stipitis показали примерно на 50% более низкие OUR и CER, чем K. marxianus (рис. 1 H ). Это различие может быть связано с наличием комплекса I дыхательной цепи у S.
cerevisiae и двух Crabtree-негативных видов, 0,42 ч -1 по сравнению с 0,44 и 0,47 ч -1 для K. marxianus и S. stipitis соответственно, в то время как при S. pombe ч -1 . Ферментативный метаболизм у Crabtree-положительных дрожжей отражался в выходе биомассы (Y sx ) около 0,1 г сухого веса (г DW)/г глюкозы и дыхательных коэффициентах (RQ) около 9. Дыхательный метаболизм Crabtree-отрицательных дрожжей вид был подтвержден значениями RQ 1.09 и 1,15 для K. marxianus и S. stipitis соответственно, что привело к выходу биомассы 0,44 и 0,58 г DW/г глюкозы соответственно. Интересно, что мы наблюдали различия в скорости поглощения кислорода (OUR) и скорости выделения углекислого газа (CER) при сравнении видов в каждой группе. Для Crabtree-негативных видов S. stipitis показали примерно на 50% более низкие OUR и CER, чем K. marxianus (рис. 1 H ). Это различие может быть связано с наличием комплекса I дыхательной цепи у S. stipitis , что приводит к более высокому соотношению фосфатов к кислороду (P/O) и, таким образом, к более высокому выходу АТФ на митохондриальный окисленный NADH. Это связано с более высоким выходом биомассы для S. stipitis , чем для K. marxianus. У видов, положительных по Crabtree, S. cerevisiae имели примерно в 2 раза более высокие OUR и CER, чем S. pombe , что свидетельствует о более низкой скорости метаболизма у S. pombe , что также отражается в более низкой скорости роста. для этого вида.
stipitis , что приводит к более высокому соотношению фосфатов к кислороду (P/O) и, таким образом, к более высокому выходу АТФ на митохондриальный окисленный NADH. Это связано с более высоким выходом биомассы для S. stipitis , чем для K. marxianus. У видов, положительных по Crabtree, S. cerevisiae имели примерно в 2 раза более высокие OUR и CER, чем S. pombe , что свидетельствует о более низкой скорости метаболизма у S. pombe , что также отражается в более низкой скорости роста. для этого вида.
Эффект Крэбтри связан с различиями в гликолитических, ППС и респираторных потоках.
Чтобы дополнительно охарактеризовать различия между Crabtree-положительными и отрицательными дрожжами, мы провели абсолютную количественную оценку протеома. Для этого мы сначала определили абсолютные концентрации белков в четырех отдельных эталонных образцах, сконструированных путем отдельного объединения трех повторов для каждого вида дрожжей, с помощью масс-спектрометрии (МС) с использованием абсолютного количественного определения на основе интенсивности (iBAQ) (20) с использованием стандарта динамического диапазона Proteomics. (UPS2) в качестве внутреннего стандарта.Абсолютные концентрации всех белков в протеомах затем определяли с помощью тандемной масс-метки (TMT) на основе (21) MS с использованием объединенных эталонных образцов в качестве внутренних эталонов. Это привело к количественной оценке 3925, 3765, 3612 и 4110 белков в S. cerevisiae , S. pombe , K. marxianus и S. stipitis соответственно (наборы данных S1).
(UPS2) в качестве внутреннего стандарта.Абсолютные концентрации всех белков в протеомах затем определяли с помощью тандемной масс-метки (TMT) на основе (21) MS с использованием объединенных эталонных образцов в качестве внутренних эталонов. Это привело к количественной оценке 3925, 3765, 3612 и 4110 белков в S. cerevisiae , S. pombe , K. marxianus и S. stipitis соответственно (наборы данных S1).
Мы приступили к изучению метаболических адаптаций и механизмов, лежащих в основе эффекта Крэбтри, с использованием GEM с ограничениями по ферментам (ec) (9).Этот метод расширяет классический подход анализа баланса потоков (FBA) (22), используемый для GEM, путем включения подробного описания ферментативных требований метаболических реакций, а также позволяет интегрировать данные об изобилии белка. Мы использовали набор инструментов GECKO (23) для создания моделей двух недавно опубликованных моделей ec для S. cerevisiae и K. marxianus с учетом экспериментально измеренных обменных потоков, скорости роста и уровней белка и использовали моделей для прогнозирования распределения потока в этих условиях (рис. 2 А ). Исследование на основе моделей было ограничено этими двумя моделями из-за их качества и используемых стандартизированных форматов, что облегчило сравнение результатов моделирования (24). В распределении потока наблюдались два основных отличия. Во-первых, мы наблюдали разницу в потоке, поступающем в ПФП от глюкозо-6-фосфата (Г6Ф). Хотя абсолютный поток через PPP у S. cerevisiae был выше, 1,25 по сравнению с 0,9 ммоль/гВт/ч, относительная доля G6P, исключая потерю G6P с полисахаридными компонентами биомассы через глюкозо-1-фосфат, метаболизировалась в ППС составил 7% по сравнению с 20.5% в К. marxianus . Это согласуется с ранее наблюдаемыми различиями в распределении потоков между Crabtree-положительными и -отрицательными дрожжами, включая S. stipitis (25⇓-27). Во-вторых, мы наблюдали большую разницу в метаболизме пирувата. S. cerevisiae имел более чем в три раза более высокий поток через гликолиз, чем K. marxianus , и большая часть пирувата потреблялась пируватдекарбоксилазой (PDC), после чего производилась продукция этанола и ацетата алкоголь- и альдегиддегидрогеназами ( АДГ и АЛД).
2 А ). Исследование на основе моделей было ограничено этими двумя моделями из-за их качества и используемых стандартизированных форматов, что облегчило сравнение результатов моделирования (24). В распределении потока наблюдались два основных отличия. Во-первых, мы наблюдали разницу в потоке, поступающем в ПФП от глюкозо-6-фосфата (Г6Ф). Хотя абсолютный поток через PPP у S. cerevisiae был выше, 1,25 по сравнению с 0,9 ммоль/гВт/ч, относительная доля G6P, исключая потерю G6P с полисахаридными компонентами биомассы через глюкозо-1-фосфат, метаболизировалась в ППС составил 7% по сравнению с 20.5% в К. marxianus . Это согласуется с ранее наблюдаемыми различиями в распределении потоков между Crabtree-положительными и -отрицательными дрожжами, включая S. stipitis (25⇓-27). Во-вторых, мы наблюдали большую разницу в метаболизме пирувата. S. cerevisiae имел более чем в три раза более высокий поток через гликолиз, чем K. marxianus , и большая часть пирувата потреблялась пируватдекарбоксилазой (PDC), после чего производилась продукция этанола и ацетата алкоголь- и альдегиддегидрогеназами ( АДГ и АЛД). Лишь небольшая часть пирувата метаболизируется пируватдегидрогеназой (ПДГ) и поступает в цикл ТСА. У K. marxianus большая часть пирувата метаболизируется ПДГ и поступает в цикл ТСА. Это указывает на то, что цикл TCA играет двойную роль у Crabtree-негативных дрожжей, выступая как в качестве источника метаболитов-предшественников, так и в дыхательном метаболизме пирувата. У S. cerevisiae основными реакциями переноса потока были ранние этапы цикла, что указывает на то, что его основная роль во время быстрого роста заключается в обеспечении предшественников компонентов биомассы посредством синтеза ɑ-кетоглутарата.Различия в метаболизме пирувата отражались в уровнях белков вокруг точки разветвления пирувата ( SI Приложение , рис. S3). Мы наблюдали гораздо большее количество ферментов, участвующих в брожении, включая PDC, ADH и ALD у Crabtree-положительных дрожжей, в то время как Crabtree-отрицательные дрожжи имели более высокие уровни PDH. У S. cerevisiae PDC обладает более высокой активностью, чем PDH, и поэтому предполагается, что он является важным фактором, управляющим ферментацией при высоком гликолитическом потоке (28).
Лишь небольшая часть пирувата метаболизируется пируватдегидрогеназой (ПДГ) и поступает в цикл ТСА. У K. marxianus большая часть пирувата метаболизируется ПДГ и поступает в цикл ТСА. Это указывает на то, что цикл TCA играет двойную роль у Crabtree-негативных дрожжей, выступая как в качестве источника метаболитов-предшественников, так и в дыхательном метаболизме пирувата. У S. cerevisiae основными реакциями переноса потока были ранние этапы цикла, что указывает на то, что его основная роль во время быстрого роста заключается в обеспечении предшественников компонентов биомассы посредством синтеза ɑ-кетоглутарата.Различия в метаболизме пирувата отражались в уровнях белков вокруг точки разветвления пирувата ( SI Приложение , рис. S3). Мы наблюдали гораздо большее количество ферментов, участвующих в брожении, включая PDC, ADH и ALD у Crabtree-положительных дрожжей, в то время как Crabtree-отрицательные дрожжи имели более высокие уровни PDH. У S. cerevisiae PDC обладает более высокой активностью, чем PDH, и поэтому предполагается, что он является важным фактором, управляющим ферментацией при высоком гликолитическом потоке (28). Наши данные о GUR, уровнях белка вокруг точки разветвления пирувата предполагают, что при более низком гликолитическом потоке у Crabtree-негативных дрожжей способность PDH достаточна для поддержания полной дыхательной диссимиляции пирувата, что указывает на то, что балансирование гликолитического потока является важным фактором. аспект в эффекте Крэбтри.
Наши данные о GUR, уровнях белка вокруг точки разветвления пирувата предполагают, что при более низком гликолитическом потоке у Crabtree-негативных дрожжей способность PDH достаточна для поддержания полной дыхательной диссимиляции пирувата, что указывает на то, что балансирование гликолитического потока является важным фактором. аспект в эффекте Крэбтри.
Адаптации в метаболизме и распределении протеома отражают компромисс между использованием глюкозы и выходом АТФ. ( A ) Распределение потоков центрального углеродного метаболизма, предсказанное FBA с использованием моделей с ограничениями по ферментам, ограниченных экспериментально измеренными обменными потоками и уровнями белка.( B ) Суммарное распределение протеома по выбранным процессам в зависимости от GUR. Белковые члены каждого процесса были определены аннотацией GO. Показаны индивидуальные значения и средние значения биологических трипликатов. Цветные прямоугольники представляют организм, как показано на рис. 1 F . См. Приложение SI , рис. S3 для абсолютного количества путей. ( C ) В этом исследовании измерено количество белков переносчиков глюкозы. Показаны средние значения ± стандартное отклонение биологических троек.( D ) Производительность АТФ рассчитана на основе результатов моделирования FBA. ( E ) Выход АТФ, рассчитанный по результатам моделирования FBA. Spo, S. pombe ; Sce, S. cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis .
1 F . См. Приложение SI , рис. S3 для абсолютного количества путей. ( C ) В этом исследовании измерено количество белков переносчиков глюкозы. Показаны средние значения ± стандартное отклонение биологических троек.( D ) Производительность АТФ рассчитана на основе результатов моделирования FBA. ( E ) Выход АТФ, рассчитанный по результатам моделирования FBA. Spo, S. pombe ; Sce, S. cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis .
Распределение протеома отражает компромисс между стратегиями использования глюкозы и выходом АТФ.
Затем мы проанализировали данные протеомики, чтобы оценить, как распределение потока отражается в распределении протеома.Суммируя содержание отдельных белков на основе аннотации термина GO (29), мы наблюдали различия в распределении белков в митохондриях и гликолизе между Crabtree-позитивными и -негативными дрожжами, в то время как пути биосинтеза PPP и аминокислот показали сходное распределение, сравнивая две группы (рис. 2 B ). Кроме того, мы не наблюдали четких различий в распределении или изобилии для всех других основных процессов биосинтеза ( SI, Приложение , рис. S4), что указывает на то, что основные метаболические различия заключаются в энергетическом метаболизме.У Crabtree-позитивных видов, у которых поток гликолиза был высоким, мы наблюдали более высокое распределение белков в направлении гликолиза за счет распределения белков в направлении митохондрий. Аналогичные тенденции также наблюдались в абсолютных уровнях белка в терминологических группах GO ( SI, Приложение , рис. S5), за исключением S. cerevisiae , показывающего более высокое суммарное содержание гликолитических белков, чем S. pombe , возможно, как в результате более высокого содержания белка (рис. 1 G ).Это открытие указывает на то, что размер протеома является важным фактором, определяющим скорость роста Crabtree-позитивных дрожжей.
2 B ). Кроме того, мы не наблюдали четких различий в распределении или изобилии для всех других основных процессов биосинтеза ( SI, Приложение , рис. S4), что указывает на то, что основные метаболические различия заключаются в энергетическом метаболизме.У Crabtree-позитивных видов, у которых поток гликолиза был высоким, мы наблюдали более высокое распределение белков в направлении гликолиза за счет распределения белков в направлении митохондрий. Аналогичные тенденции также наблюдались в абсолютных уровнях белка в терминологических группах GO ( SI, Приложение , рис. S5), за исключением S. cerevisiae , показывающего более высокое суммарное содержание гликолитических белков, чем S. pombe , возможно, как в результате более высокого содержания белка (рис. 1 G ).Это открытие указывает на то, что размер протеома является важным фактором, определяющим скорость роста Crabtree-позитивных дрожжей.
Важным событием в эволюции S. cerevisiae стала полногеномная дупликация (WGD) (30, 31), которая привела к увеличению количества генов гликолитических белков (32), что считается основным фактором Эффект Крэбтри. Действительно, мы наблюдали более высокие уровни всех гликолитических белков у обоих Крэбтри-положительных видов по сравнению с Крэбтри-отрицательными ( SI Приложение , рис.С6). WGD не был обнаружен у S. pombe , и, таким образом, дрожжи полагались на мелкомасштабные события дупликации, а также на увеличение экспрессии отдельных ферментов (33). Интересно, что хотя S. pombe имеет меньше дуплицированных гликолитических генов, чем S. cerevisiae , моноизоформные белки имеют такое же количество, что и совокупное количество паралогов S. cerevisiae , особенно в верхней половине гликолиза. Кроме того, несколько исследований показали, что транспорт глюкозы является основной стадией гликолиза, контролирующей поток, и, таким образом, служит одним из основных факторов, влияющих на эффект Крэбтри (19, 32, 34).
cerevisiae стала полногеномная дупликация (WGD) (30, 31), которая привела к увеличению количества генов гликолитических белков (32), что считается основным фактором Эффект Крэбтри. Действительно, мы наблюдали более высокие уровни всех гликолитических белков у обоих Крэбтри-положительных видов по сравнению с Крэбтри-отрицательными ( SI Приложение , рис.С6). WGD не был обнаружен у S. pombe , и, таким образом, дрожжи полагались на мелкомасштабные события дупликации, а также на увеличение экспрессии отдельных ферментов (33). Интересно, что хотя S. pombe имеет меньше дуплицированных гликолитических генов, чем S. cerevisiae , моноизоформные белки имеют такое же количество, что и совокупное количество паралогов S. cerevisiae , особенно в верхней половине гликолиза. Кроме того, несколько исследований показали, что транспорт глюкозы является основной стадией гликолиза, контролирующей поток, и, таким образом, служит одним из основных факторов, влияющих на эффект Крэбтри (19, 32, 34). Это дополнительно подтверждается обнаружением того, что эффект Крэбтри устранен у штамма S. cerevisiae с ограниченной способностью поглощения глюкозы (35), но это также сопровождается более медленным ростом. Помимо различий в GUR, наши данные по переносчикам гексозы (HXT) показывают ключевые различия как в уровнях, так и в количестве активных переносчиков глюкозы (рис. 2 C ). Два Крэбтри-негативных штамма, судя по распространенности, полагаются в основном на один переносчик глюкозы, основное различие заключается в том, что 90–155 K.marxianus экспрессирует низкоаффинный транспортер RAG1, тогда как S. stipitis экспрессирует высокоаффинный транспортер HGT2. Оба Crabtree-положительные дрожжи обладают дополнительными изоформами переносчиков глюкозы, всего 17, из которых 6 экспрессируются в нормальных условиях в случае S. cerevisiae и 8 в случае S. pombe , что подчеркивает важность транспорт глюкозы в эффекте Крэбтри (34). Однако эти два вида различаются по распределению экспрессии транспортеров в изучаемых условиях.
Это дополнительно подтверждается обнаружением того, что эффект Крэбтри устранен у штамма S. cerevisiae с ограниченной способностью поглощения глюкозы (35), но это также сопровождается более медленным ростом. Помимо различий в GUR, наши данные по переносчикам гексозы (HXT) показывают ключевые различия как в уровнях, так и в количестве активных переносчиков глюкозы (рис. 2 C ). Два Крэбтри-негативных штамма, судя по распространенности, полагаются в основном на один переносчик глюкозы, основное различие заключается в том, что 90–155 K.marxianus экспрессирует низкоаффинный транспортер RAG1, тогда как S. stipitis экспрессирует высокоаффинный транспортер HGT2. Оба Crabtree-положительные дрожжи обладают дополнительными изоформами переносчиков глюкозы, всего 17, из которых 6 экспрессируются в нормальных условиях в случае S. cerevisiae и 8 в случае S. pombe , что подчеркивает важность транспорт глюкозы в эффекте Крэбтри (34). Однако эти два вида различаются по распределению экспрессии транспортеров в изучаемых условиях. S. cerevisiae распределяет экспрессию между несколькими переносчиками с разным сродством к глюкозе, в то время как S. pombe преимущественно имеет высокий уровень экспрессии своего высокоаффинного транспортера ght5.
S. cerevisiae распределяет экспрессию между несколькими переносчиками с разным сродством к глюкозе, в то время как S. pombe преимущественно имеет высокий уровень экспрессии своего высокоаффинного транспортера ght5.
Различия в стратегиях метаболизма глюкозы дополнительно отражаются в моделировании производства АТФ. Чистый поток продукции АТФ, рассчитанный путем объединения продукции АТФ в результате гликолиза и дыхания и равный потреблению АТФ клеткой, составил 36.5 и 30,5 ммоль/гВт/ч для S. cerevisiae и K. marxianus соответственно (рис. 2 D ). Однако выход АТФ, определенный в ммоль АТФ/ммоль глюкозы, был в три раза выше у K. marxianus (рис. 2 E ), что отражает более высокую эффективность дыхательного метаболизма по выходу АТФ. Эти результаты ясно подчеркивают разницу в двух стратегиях, используемых для производства энергии. Крэбтри-положительные дрожжи производят АТФ неэффективным путем ферментации и компенсируют низкую эффективность за счет большого поглощения глюкозы и потока посредством гликолиза, в то время как Крэбтри-отрицательные дрожжи уравновешивают поток посредством гликолиза и дыхания, что приводит к высокому выходу АТФ.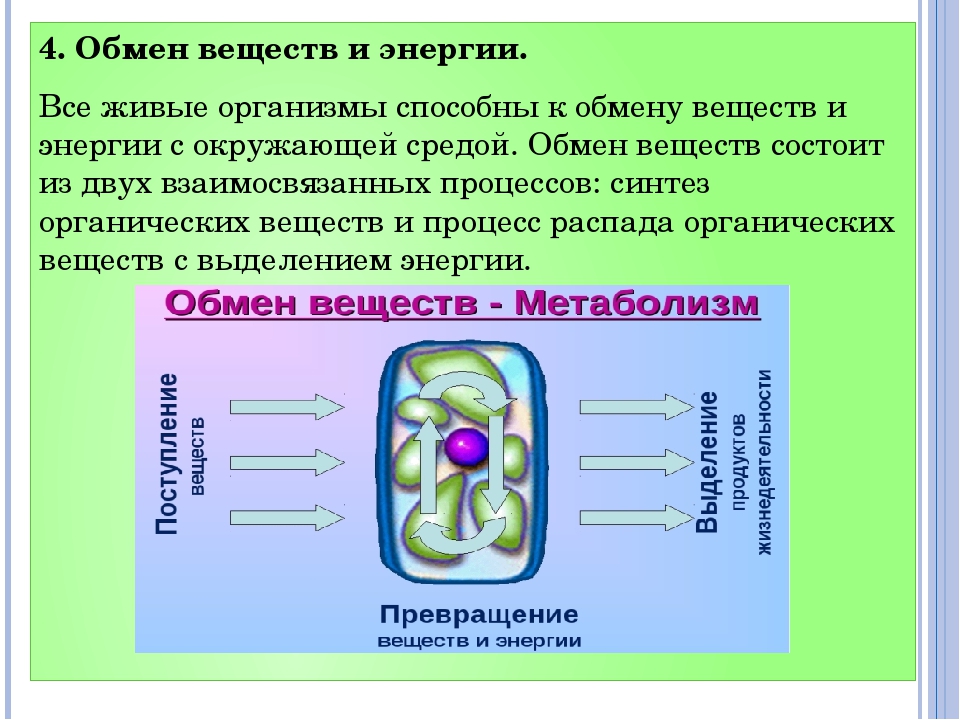 Кроме того, K. marxianus более эффективно использует АТФ для производства биомассы, чем S. cerevisiae , и это также может способствовать способности к быстрому росту без избыточного метаболизма для Крэбтри-негативных дрожжей.
Кроме того, K. marxianus более эффективно использует АТФ для производства биомассы, чем S. cerevisiae , и это также может способствовать способности к быстрому росту без избыточного метаболизма для Крэбтри-негативных дрожжей.
Ограничения в ETC и АТФ-синтазе характерны для эффекта Крэбтри.
Чтобы дополнительно охарактеризовать адаптацию, связанную с эффектом Крэбтри, мы рассчитали использование емкости, определяемое как отношение между измеренным содержанием фермента и потребностью в ферменте, как предсказано моделями ec S.cerevisiae и К. marxianus . Мы наблюдали более высокое использование в гликолизе у S. cerevisiae , в то время как K. marxianus имели более высокое использование в цикле TCA, в соответствии с различиями, наблюдаемыми на уровне потока (рис. 3 A ). Это означает, что усилению потоков через любой из этих путей способствует не только повышенная экспрессия, но и более высокая занятость составляющих их ферментов. Кроме того, для обоих дрожжей мы наблюдали полное использование белков ЭТЦ и АТФ-синтазы.Это разумно, поскольку все эти белки являются митохондриальными и присутствуют во внутренней мембране митохондрий, и любой неиспользованный белок в этих комплексах приведет к неэффективному использованию пространства митохондриальной мембраны. Мы также наблюдали разницу в использовании ферментов в PPP, с более низким использованием в K. marxianus , возможно, в результате более низкого абсолютного потока через PPP. Однако при рассмотрении абсолютных уровней белка четкой разницы не наблюдалось ( SI Приложение , рис.С7).
Кроме того, для обоих дрожжей мы наблюдали полное использование белков ЭТЦ и АТФ-синтазы.Это разумно, поскольку все эти белки являются митохондриальными и присутствуют во внутренней мембране митохондрий, и любой неиспользованный белок в этих комплексах приведет к неэффективному использованию пространства митохондриальной мембраны. Мы также наблюдали разницу в использовании ферментов в PPP, с более низким использованием в K. marxianus , возможно, в результате более низкого абсолютного потока через PPP. Однако при рассмотрении абсолютных уровней белка четкой разницы не наблюдалось ( SI Приложение , рис.С7).
Ограничения в ЭТЦ и АТФ-синтазе характерны для эффекта Крэбтри. ( A ) Использование мощности, рассчитанное как предсказанные моделью уровни ферментов, деленные на экспериментально измеренные уровни ферментов, основных путей центрального углеродного метаболизма (CCM). ( B ) Суммарное распределение протеома по процессам CCM в зависимости от GUR. Белковые члены каждого процесса были определены аннотацией GO. Показаны средние значения биологических троек.Цветные прямоугольники представляют организм, как указано на панели C . См. Приложение SI , рис. S7 A для абсолютного содержания процессов. ( C ) Абсолютное содержание белка в компонентах, составляющих ЭТЦ и АТФ-синтазу. Показаны средние значения ± стандартное отклонение биологических троек. См. SI Приложение , рис. S7 B для распределения компонентов протеома. PPP, пентозофосфатный путь; Spo, S. pombe ; Сце, С.cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis .
Белковые члены каждого процесса были определены аннотацией GO. Показаны средние значения биологических троек.Цветные прямоугольники представляют организм, как указано на панели C . См. Приложение SI , рис. S7 A для абсолютного содержания процессов. ( C ) Абсолютное содержание белка в компонентах, составляющих ЭТЦ и АТФ-синтазу. Показаны средние значения ± стандартное отклонение биологических троек. См. SI Приложение , рис. S7 B для распределения компонентов протеома. PPP, пентозофосфатный путь; Spo, S. pombe ; Сце, С.cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis .
Затем мы проанализировали распределение протеома в митохондриальном энергетическом метаболизме, включая цикл TCA, ETC и АТФ-синтазу, у четырех дрожжей (рис. 3 B ). Здесь мы наблюдали более высокое распределение всех трех процессов у Crabtree-негативных дрожжей, где распределение циклу ТСА и АТФ-синтазе составляло около 3%, в то время как около 1,5% приходилось на ЭТЦ по сравнению с примерно 0,5 и 0. .25% протеома у Crabtree-положительных дрожжей. Эти результаты также были отражены в абсолютных уровнях содержания отростков и отдельных ферментных комплексов (рис. 3 C и SI Приложение , рис. S8 и S9). Для всех компонентов ETC и АТФ-синтазы мы наблюдали примерно в три раза более высокое содержание у Crabtree-негативных дрожжей по сравнению с Crabtree-позитивными дрожжами. Это ясно подчеркивает разницу в стратегии, используемой для производства АТФ. В совокупности с более высоким выделением гликолитических белков за счет митохондриальных у Crabtree-позитивных дрожжей это свидетельствует о том, что высокий гликолитический поток вызывает насыщение способности окисления НАДН посредством дыхания, вызывая усиление брожения при высоких значениях GUR.Это согласуется с предыдущим исследованием S. cerevisiae , показывающим, что увеличение способности окисления НАДН за счет экспрессии альтернативной оксидазы (АОХ), что позволяет напрямую восстанавливать кислород из пула убихинонов, приводит к уменьшению образования этанола (36).
.25% протеома у Crabtree-положительных дрожжей. Эти результаты также были отражены в абсолютных уровнях содержания отростков и отдельных ферментных комплексов (рис. 3 C и SI Приложение , рис. S8 и S9). Для всех компонентов ETC и АТФ-синтазы мы наблюдали примерно в три раза более высокое содержание у Crabtree-негативных дрожжей по сравнению с Crabtree-позитивными дрожжами. Это ясно подчеркивает разницу в стратегии, используемой для производства АТФ. В совокупности с более высоким выделением гликолитических белков за счет митохондриальных у Crabtree-позитивных дрожжей это свидетельствует о том, что высокий гликолитический поток вызывает насыщение способности окисления НАДН посредством дыхания, вызывая усиление брожения при высоких значениях GUR.Это согласуется с предыдущим исследованием S. cerevisiae , показывающим, что увеличение способности окисления НАДН за счет экспрессии альтернативной оксидазы (АОХ), что позволяет напрямую восстанавливать кислород из пула убихинонов, приводит к уменьшению образования этанола (36). ). Интересно, что S. stipitis изначально обладает AOX, STO1. Поэтому, чтобы исследовать роль АО, мы сравнили содержание Sto1p с содержанием комплекса IV (оксидаза цитохрома c ). Мы наблюдали пятикратное увеличение медианы содержания субъединиц комплекса IV ( SI Приложение , рис.S10), что указывает на то, что основной стадией потребления кислорода является оксидаза цитохрома c и что АОХ используется в меньшей степени в тестируемых условиях. Кроме того, в S. stipitis присутствует комплекс I. Мы наблюдали экспрессию комплекса, но не внутренней NADH-дегидрогеназы, NDI1, предполагая, что окисление NADH зависит от комплекса I и внешней NADH-дегидрогеназы, NDE1, в изучаемых условиях. Присутствие комплекса I представляет собой дополнительную стадию переноса протонов, что, в свою очередь, приводит к более высокому выходу P/O и АТФ.Это может объяснить значительно более высокий выход биомассы, а также более низкие OUR и CER у S. stipitis по сравнению с K.
). Интересно, что S. stipitis изначально обладает AOX, STO1. Поэтому, чтобы исследовать роль АО, мы сравнили содержание Sto1p с содержанием комплекса IV (оксидаза цитохрома c ). Мы наблюдали пятикратное увеличение медианы содержания субъединиц комплекса IV ( SI Приложение , рис.S10), что указывает на то, что основной стадией потребления кислорода является оксидаза цитохрома c и что АОХ используется в меньшей степени в тестируемых условиях. Кроме того, в S. stipitis присутствует комплекс I. Мы наблюдали экспрессию комплекса, но не внутренней NADH-дегидрогеназы, NDI1, предполагая, что окисление NADH зависит от комплекса I и внешней NADH-дегидрогеназы, NDE1, в изучаемых условиях. Присутствие комплекса I представляет собой дополнительную стадию переноса протонов, что, в свою очередь, приводит к более высокому выходу P/O и АТФ.Это может объяснить значительно более высокий выход биомассы, а также более низкие OUR и CER у S. stipitis по сравнению с K. marxianus (рис. 1 G и H ).
marxianus (рис. 1 G и H ).
Эффект Крэбтри сопровождается различиями в содержании РП и эффективности трансляции белков.
Эффект Крэбтри развился независимо во многих линиях дрожжей, что свидетельствует о эволюционном преимуществе перехода к ферментации при избытке глюкозы.Считается, что это преимущество заключается в более высокой скорости производства АТФ и более низкой стоимости протеома, что позволяет Crabtree-положительным дрожжам расти быстрее (37). Однако наши данные показывают, что Crabtree-негативные дрожжи K. marxianus и S. stipitis растут с той же скоростью, что и Crabtree-положительные дрожжи S. cerevisiae , в то время как Crabtree-положительные дрожжи S. pombe значительно более низкая скорость роста (рис. 1 G ). Это повышает вероятность того, что в условиях избытка глюкозы Crabtree-позитивные дрожжи подвергаются дополнительным физиологическим ограничениям, что уравновешивается преимуществом, которое дает высокая скорость продукции АТФ посредством ферментации глюкозы. Одним из основных кандидатов на такое ограничение является трансляция белка. Это очевидно с эволюционной точки зрения: основным эволюционным событием в линии Saccharomyces , которое имеет ярко выраженный эффект Крэбтри, является событие WGD, за которым следует массовая потеря генов, что приводит к сохранению около 10% дуплицированных генов, которые эволюционировали. на паралоги (38). Две основные группы сохраненных паралогов: 1) ферменты углеводного обмена, которые обеспечивают более высокий гликолитический поток и, таким образом, вызывают выраженный эффект Крэбтри у дрожжей после WGD; и 2) РП (39).У S. cerevisiae 59 из 78 субъединиц RP имеют паралоги ( SI Приложение , рис. S11 A и B ). В S. pombe , который не принадлежит к линии Saccharomyces , но независимо эволюционировал и стал положительным по Крэбтри, 62 из 78 субъединиц RP имеют паралоги ( SI Приложение , рис. S11 A и B ). Напротив, K.
Одним из основных кандидатов на такое ограничение является трансляция белка. Это очевидно с эволюционной точки зрения: основным эволюционным событием в линии Saccharomyces , которое имеет ярко выраженный эффект Крэбтри, является событие WGD, за которым следует массовая потеря генов, что приводит к сохранению около 10% дуплицированных генов, которые эволюционировали. на паралоги (38). Две основные группы сохраненных паралогов: 1) ферменты углеводного обмена, которые обеспечивают более высокий гликолитический поток и, таким образом, вызывают выраженный эффект Крэбтри у дрожжей после WGD; и 2) РП (39).У S. cerevisiae 59 из 78 субъединиц RP имеют паралоги ( SI Приложение , рис. S11 A и B ). В S. pombe , который не принадлежит к линии Saccharomyces , но независимо эволюционировал и стал положительным по Крэбтри, 62 из 78 субъединиц RP имеют паралоги ( SI Приложение , рис. S11 A и B ). Напротив, K. marxianus и S. stipitis не имеют паралогов RP ( SI Приложение , рис.S11 A и B ). Эволюция и сохранение паралогов RP у Crabtree-позитивных дрожжей предполагает, что рибосомы у этих видов могли стать более специализированными (например, транслировать разные субнаборы белков), что может сопровождаться более медленной кинетикой трансляции белков. Поэтому мы сравнили содержание и эффективность рибосом у Crabtree-положительных и -отрицательных дрожжей.
marxianus и S. stipitis не имеют паралогов RP ( SI Приложение , рис.S11 A и B ). Эволюция и сохранение паралогов RP у Crabtree-позитивных дрожжей предполагает, что рибосомы у этих видов могли стать более специализированными (например, транслировать разные субнаборы белков), что может сопровождаться более медленной кинетикой трансляции белков. Поэтому мы сравнили содержание и эффективность рибосом у Crabtree-положительных и -отрицательных дрожжей.
В наших протеомных данных мы обнаружили от 74 до 77% RP в S. cerevisiae и S.pombe соответственно, в то время как для K. marxianus и S. stipitis было обнаружено от 97 до 98% RP ( SI Приложение , рис. S11 A ). Однако у всех видов мы обнаружили от 96 до 99% субъединиц RP, при этом субъединица считается обнаруженной, если обнаруживается один из двух паралогов ( SI Приложение , рис. S11 B ). Общее содержание РФП составляло от 19,5 до 23,6% (г на г общего белка) у быстрорастущих видов ( S. cerevisiae , K.marxianus и S. stipitis ) и 16,1% у более медленно растущего S. pombe ( SI Приложение , рис. S11 C ), в соответствии с предыдущими исследованиями, показывающими, что количество RP увеличивается линейно с удельная скорость роста (40, 41). Затем мы рассчитали общую эффективность рибосомной трансляции как мг общего белка на нмоль рибосом в час и обнаружили, что S. pombe имеет менее эффективные рибосомы по сравнению с S. stipitis , тогда как K.marxianus и S. cerevisiae показали сходную эффективность (рис. 4 A ). Это указывает на то, что эффективность трансляции является фактором, лежащим в основе эффекта Крэбтри, хотя для подтверждения этого потребуются данные о большем количестве видов дрожжей. Поскольку мы показали, что ETC и АТФ-синтаза особенно ограничивают центральный углеродный метаболизм и выработку энергии (рис. 3 A ), мы затем изучили количество митохондриальных рибосомных белков (MRP) в нашем наборе данных.
cerevisiae , K.marxianus и S. stipitis ) и 16,1% у более медленно растущего S. pombe ( SI Приложение , рис. S11 C ), в соответствии с предыдущими исследованиями, показывающими, что количество RP увеличивается линейно с удельная скорость роста (40, 41). Затем мы рассчитали общую эффективность рибосомной трансляции как мг общего белка на нмоль рибосом в час и обнаружили, что S. pombe имеет менее эффективные рибосомы по сравнению с S. stipitis , тогда как K.marxianus и S. cerevisiae показали сходную эффективность (рис. 4 A ). Это указывает на то, что эффективность трансляции является фактором, лежащим в основе эффекта Крэбтри, хотя для подтверждения этого потребуются данные о большем количестве видов дрожжей. Поскольку мы показали, что ETC и АТФ-синтаза особенно ограничивают центральный углеродный метаболизм и выработку энергии (рис. 3 A ), мы затем изучили количество митохондриальных рибосомных белков (MRP) в нашем наборе данных. Количество MRP у четырех исследованных видов дрожжей колеблется от 66 до 77 субъединиц, и у всех четырех видов мы обнаружили от 90 до 99% этих белков ( SI Приложение , рис. S11 D ). Здесь мы обнаруживаем, что S. cerevisiae и S. pombe экспрессировали MRP только на 28% от уровней экспрессии MRP, обнаруженных в K. marxianus и S. stipitis (рис. 4 B ). Поскольку MRP транслируют ключевые субъединицы АТФ-синтазы, оксидазы цитохрома c и миторибосомы, мы рассчитали эффективность трансляции миторибосомных белков (мг белков, транслируемых миторибосомами, на нмоль миторибосом в час), основываясь на среднем молярном содержании всех субъединиц этих субъединиц. три комплекса.Наши данные показали, что эффективность трансляции миторибосом у S. pombe была намного ниже, чем у S. stipitis , в то время как эффективность K. marxianus и S. cerevisiae была сходной (рис. 4 C). ).
Количество MRP у четырех исследованных видов дрожжей колеблется от 66 до 77 субъединиц, и у всех четырех видов мы обнаружили от 90 до 99% этих белков ( SI Приложение , рис. S11 D ). Здесь мы обнаруживаем, что S. cerevisiae и S. pombe экспрессировали MRP только на 28% от уровней экспрессии MRP, обнаруженных в K. marxianus и S. stipitis (рис. 4 B ). Поскольку MRP транслируют ключевые субъединицы АТФ-синтазы, оксидазы цитохрома c и миторибосомы, мы рассчитали эффективность трансляции миторибосомных белков (мг белков, транслируемых миторибосомами, на нмоль миторибосом в час), основываясь на среднем молярном содержании всех субъединиц этих субъединиц. три комплекса.Наши данные показали, что эффективность трансляции миторибосом у S. pombe была намного ниже, чем у S. stipitis , в то время как эффективность K. marxianus и S. cerevisiae была сходной (рис. 4 C). ). Эти три фактора — менее эффективные цитоплазматические рибосомы, более низкая экспрессия миторибосом и/или менее эффективные миторибосомы — вместе приводят к значительно более низкой экспрессии АТФ-синтазы и цитохром c оксидазы как у S.cerevisiae и S. pombe , при 17% уровней экспрессии, обнаруженных у K. marxianus и S. stipitis (рис. 4 D ). Таким образом, в совокупности наши данные показывают, что эффект Крэбтри у дрожжей сопровождается адаптацией метаболизма, особенно стратегий, используемых для производства АТФ, а также адаптацией трансляции белков, эффективности и/или содержания рибосом и миторибосом.
Эти три фактора — менее эффективные цитоплазматические рибосомы, более низкая экспрессия миторибосом и/или менее эффективные миторибосомы — вместе приводят к значительно более низкой экспрессии АТФ-синтазы и цитохром c оксидазы как у S.cerevisiae и S. pombe , при 17% уровней экспрессии, обнаруженных у K. marxianus и S. stipitis (рис. 4 D ). Таким образом, в совокупности наши данные показывают, что эффект Крэбтри у дрожжей сопровождается адаптацией метаболизма, особенно стратегий, используемых для производства АТФ, а также адаптацией трансляции белков, эффективности и/или содержания рибосом и миторибосом.
Эффект Крэбтри сопровождается различиями в содержании РП и эффективности трансляции белков.( A ) Сравнение общей эффективности рибосомной трансляции, рассчитанной как миллиграммы общего белка на наномоль рибосом в час. ( B ) Сравнение уровней экспрессии MRP, выраженных в процентах от общего протеома. ( C ) Сравнение эффективности трансляции миторибосомных белков, рассчитанной на основе среднего молярного содержания всех субъединиц митохондриально экспрессируемых белковых комплексов, включая цитохром c оксидазу, АТФ-синтазу и миторибосомы.( D ) Сравнение уровней экспрессии цитохром с оксидазы и АТФ-синтазы, выраженное в процентах от общего клеточного белка. С, Спо, С. pombe ; Sce, S. cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis .
( C ) Сравнение эффективности трансляции миторибосомных белков, рассчитанной на основе среднего молярного содержания всех субъединиц митохондриально экспрессируемых белковых комплексов, включая цитохром c оксидазу, АТФ-синтазу и миторибосомы.( D ) Сравнение уровней экспрессии цитохром с оксидазы и АТФ-синтазы, выраженное в процентах от общего клеточного белка. С, Спо, С. pombe ; Sce, S. cerevisiae ; Кма, К. marxianus ; Sstip, S. Stipitis .
Обсуждение
Здесь мы количественно описали метаболические и протеомные адаптации, лежащие в основе эффекта Крэбтри, путем сравнения двух Крэбтри-положительных дрожжей, S. cerevisiae и S.pombe и два Crabtree-негативных дрожжей, K. marxianus и S. stipitis , культивируемых в биореакторах в условиях избытка глюкозы. Комбинируя физиологическую и протеомную количественную оценку с GEM, мы обнаружили, что эффект Крэбтри связан с адаптацией метаболических стратегий утилизации глюкозы, а также с различиями в организации и эффективности как митохондриальной, так и цитоплазматической трансляции.
Оптимальное использование углерода имеет решающее значение для производства энергии в форме АТФ и предшественников биомассы для роста клеток.Крэбтри-положительные дрожжи производят АТФ в основном за счет использования гликолиза и ферментации, что приводит к образованию побочных продуктов, в то время как Крэбтри-отрицательные дрожжи полностью окисляют глюкозу до углекислого газа и воды. Хотя полное окисление глюкозы более эффективно с точки зрения образования АТФ на молекулу глюкозы (3), оно достигается за счет высокой стоимости белка. С другой стороны, метаболизм глюкозы посредством ферментации сопряжен с низким выходом АТФ, что потенциально приводит к более низкой скорости роста, но имеет более высокую эффективность с точки зрения стоимости белка (1).Чтобы компенсировать низкий выход, потребуется повышенный гликолитический поток. Наши данные подтверждают, что Crabtree-положительные дрожжи действительно имеют более высокую скорость поглощения глюкозы и гликолиза. Мы наблюдали примерно вдвое и втрое более высокую скорость поглощения у S. pombe и S. cerevisiae соответственно, а также более высокий гликолитический поток по сравнению с Crabtree-негативными дрожжами. Моделирование также показывает, что потоки вокруг точки разветвления пирувата значительно различаются, где S.cerevisiae продемонстрировал высокий поток через PDC и образование этанола, в то время как K. marxianus показал более высокий поток через цикл TCA и дыхание. Хотя общий АТФ был одинаковым у двух дрожжей, молярный выход АТФ на глюкозу был почти в три раза выше у K. marxianus . Это ясно подчеркивает компромисс в производстве АТФ с использованием различных метаболических стратегий.
pombe и S. cerevisiae соответственно, а также более высокий гликолитический поток по сравнению с Crabtree-негативными дрожжами. Моделирование также показывает, что потоки вокруг точки разветвления пирувата значительно различаются, где S.cerevisiae продемонстрировал высокий поток через PDC и образование этанола, в то время как K. marxianus показал более высокий поток через цикл TCA и дыхание. Хотя общий АТФ был одинаковым у двух дрожжей, молярный выход АТФ на глюкозу был почти в три раза выше у K. marxianus . Это ясно подчеркивает компромисс в производстве АТФ с использованием различных метаболических стратегий.
Эта разница в метаболизме также отражается на уровне протеома.Наряду с более высоким гликолитическим потоком мы наблюдали более высокие уровни гликолитических белков у обоих Crabtree-позитивных дрожжей, в то время как уровни белка в цикле TCA, ETC и АТФ-синтазы были выше у Crabtree-негативных дрожжей. Это указывает на то, что увеличение гликолитического потока и связанное с ним увеличение уровня белка происходит за счет дыхательных белков, что также видно из наших данных, сравнивающих распределение ресурсов протеома. Это согласуется с подавлением дыхания глюкозой в результате перестройки регуляции, связанной с эволюцией аэробного брожения (42⇓–44).
Это согласуется с подавлением дыхания глюкозой в результате перестройки регуляции, связанной с эволюцией аэробного брожения (42⇓–44).
Эффект Крэбтри развился независимо у S. cerevisiae и S. pombe (13). Основным фактором развития эффекта Крэбтри у S. cerevisiae является WGD, включая дублирование 6 из 13 гликолитических ферментов и увеличение количества HXT, что приводит к увеличению гликолитического потока (32, 34). У S. pombe WGD не наблюдалось, и присутствовало меньше гликолитических паралогов (33). Интересно, что хотя приток глюкозы в 90–155 S.pombe примерно в два раза меньше, чем у S. cerevisiae , некоторые отдельные гликолитические ферменты на ранних стадиях гликолиза демонстрируют уровень, сходный с суммарным содержанием изоферментов S. cerevisiae . Ферменты на более поздних стадиях показали меньшее количество, что указывает на ограничение во второй половине гликолиза. Возможным объяснением является более низкая активность PYK, важного регулятора гликолитического потока (45), у S.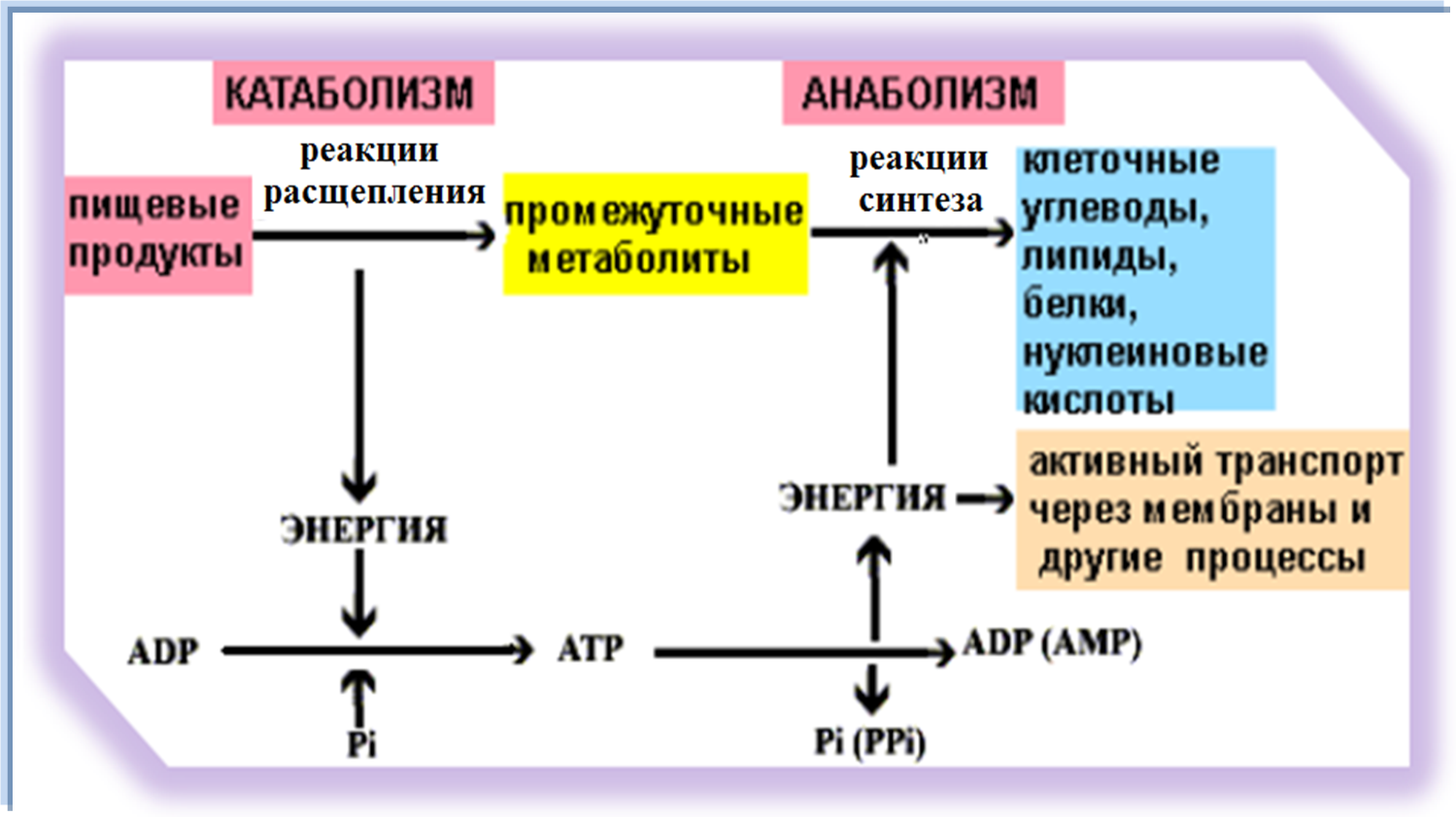 pombe в результате мутации, делающей его нечувствительным к стимуляции фруктозобисфосфатом (46).Кроме того, мы наблюдали разницу в HXT. S. cerevisiae и S. pombe содержат 17 и 8 HXT, соответственно, по сравнению с 2 у Crabtree-негативных дрожжей. S. pombe и Crabtree-негативные дрожжи экспрессировали в основном один HXT, в то время как S. cerevisiae распределяли экспрессию среди множества паралогов. Распределение потока по нескольким паралогам HXT и гликолитических белков указывает на то, что S. cerevisiae оптимизировали свой метаболизм для быстрого использования глюкозы, в то время как другие дрожжи имеют более ограниченное поглощение глюкозы.
pombe в результате мутации, делающей его нечувствительным к стимуляции фруктозобисфосфатом (46).Кроме того, мы наблюдали разницу в HXT. S. cerevisiae и S. pombe содержат 17 и 8 HXT, соответственно, по сравнению с 2 у Crabtree-негативных дрожжей. S. pombe и Crabtree-негативные дрожжи экспрессировали в основном один HXT, в то время как S. cerevisiae распределяли экспрессию среди множества паралогов. Распределение потока по нескольким паралогам HXT и гликолитических белков указывает на то, что S. cerevisiae оптимизировали свой метаболизм для быстрого использования глюкозы, в то время как другие дрожжи имеют более ограниченное поглощение глюкозы.
В дополнение к сохранению паралогичных гликолитических ферментов и HXT, Crabtree-позитивные дрожжи также эволюционировали, чтобы содержать паралоги для многих RP. Полученное в результате увеличение количества генов RP было связано с увеличением ферментативной способности (47). Гены RP сильно транскрибируются, на них приходится 50% транскрипции РНК-полимеразы II (48). Удвоение дозы генов с 76 до 77% RPs у Crabtree-позитивных дрожжей означает, что зрелые рибосомы, вероятно, будут очень разнообразными, при этом некоторые исследования оценивают, что разнообразие рибосом может превышать количество фактического содержания рибосом в клетке (49).Преимущество такого высокого разнообразия состоит в том, что он позволяет точно контролировать активность рибосом, но обязательно означает, что общая кинетика трансляции белка становится медленнее. Здесь наши анализы показывают, что цитозольная трансляционная эффективность действительно была ниже у некоторых Crabtree-позитивных видов, что усугубляло протеомные ограничения в быстрорастущих клетках и, таким образом, благоприятствовало использованию ферментативных путей для метаболизма глюкозы, когда позволяют условия. Кроме того, мы обнаружили, что выделение MRPs было значительно ниже у Crabtree-позитивных видов, что, возможно, объясняется потерей регуляторных элементов, связывающих экспрессию MRPs и RPs после WGD (50).
Удвоение дозы генов с 76 до 77% RPs у Crabtree-позитивных дрожжей означает, что зрелые рибосомы, вероятно, будут очень разнообразными, при этом некоторые исследования оценивают, что разнообразие рибосом может превышать количество фактического содержания рибосом в клетке (49).Преимущество такого высокого разнообразия состоит в том, что он позволяет точно контролировать активность рибосом, но обязательно означает, что общая кинетика трансляции белка становится медленнее. Здесь наши анализы показывают, что цитозольная трансляционная эффективность действительно была ниже у некоторых Crabtree-позитивных видов, что усугубляло протеомные ограничения в быстрорастущих клетках и, таким образом, благоприятствовало использованию ферментативных путей для метаболизма глюкозы, когда позволяют условия. Кроме того, мы обнаружили, что выделение MRPs было значительно ниже у Crabtree-позитивных видов, что, возможно, объясняется потерей регуляторных элементов, связывающих экспрессию MRPs и RPs после WGD (50). Мы также обнаружили, что эффективность митохондриальной трансляции была ниже у некоторых Crabtree-позитивных видов. Эти факторы обуславливают значительно более низкую экспрессию АТФ-синтазы и цитохром с оксидазы и подчеркивают значение адаптаций в составе и эффективности рибосом в эффекте Крэбтри.
Мы также обнаружили, что эффективность митохондриальной трансляции была ниже у некоторых Crabtree-позитивных видов. Эти факторы обуславливают значительно более низкую экспрессию АТФ-синтазы и цитохром с оксидазы и подчеркивают значение адаптаций в составе и эффективности рибосом в эффекте Крэбтри.
В заключение, это исследование дает представление о механизмах, лежащих в основе эффекта Крэбтри с точки зрения адаптации в метаболизме и трансляции белков, подтверждая, что компромисс между скоростью и выходом образования АТФ, а также рибосомной специализацией и эффективность перевода, являются важными факторами в развитии эффекта Крэбтри.Кроме того, созданные здесь наборы протеомных данных, а также наш подход к моделированию вместе обеспечивают основу для анализа данных многовидовой омики, чтобы понять процесс диверсификации видов, постоянную проблему в системах и синтетической биологии.
Методы
Условия культивирования.
В этом исследовании использовались дрожжи S. cerevisiae CEN.PK113-7D (MATa, MAL2-8c, SUC2), S. pombe (ATCC24843), K. marxianus CBS6556 и S.стебель (ATCC58376). Клетки хранили в аликвотных растворах глицерина при температуре -80 °С. Периодическое культивирование проводили в биореакторах DASGIP объемом 1 л с анализом отходящих газов, контролем pH, контролем температуры и датчиками растворенного кислорода. Использовали рабочий объем 700 мл, температуру 30 °С, рН 5, контролировали добавлением 2 М КОН и 2 М HCl, исходный инокулят с оптической плотностью 0,1. Первоначальную аэрацию и перемешивание устанавливали на 50 сл/ч -1 и 800 об/мин соответственно, а DO поддерживали выше 60% за счет увеличения скорости перемешивания.Основу всех культур составляла минимальная среда, содержащая 20 г/л глюкозы (10 г/л для K. marxianus ), 5 г/л (Nh5)2SO4, 3 г/л KH 2 PO 4 , 0,5 г/л MgSO 4 ⋅7h3O, 1 мл/л раствора микроэлементов, 1 мл/л раствора витаминов и 0,1 мл/л пеногасителя 204 (Sigma-Aldrich).
cerevisiae CEN.PK113-7D (MATa, MAL2-8c, SUC2), S. pombe (ATCC24843), K. marxianus CBS6556 и S.стебель (ATCC58376). Клетки хранили в аликвотных растворах глицерина при температуре -80 °С. Периодическое культивирование проводили в биореакторах DASGIP объемом 1 л с анализом отходящих газов, контролем pH, контролем температуры и датчиками растворенного кислорода. Использовали рабочий объем 700 мл, температуру 30 °С, рН 5, контролировали добавлением 2 М КОН и 2 М HCl, исходный инокулят с оптической плотностью 0,1. Первоначальную аэрацию и перемешивание устанавливали на 50 сл/ч -1 и 800 об/мин соответственно, а DO поддерживали выше 60% за счет увеличения скорости перемешивания.Основу всех культур составляла минимальная среда, содержащая 20 г/л глюкозы (10 г/л для K. marxianus ), 5 г/л (Nh5)2SO4, 3 г/л KH 2 PO 4 , 0,5 г/л MgSO 4 ⋅7h3O, 1 мл/л раствора микроэлементов, 1 мл/л раствора витаминов и 0,1 мл/л пеногасителя 204 (Sigma-Aldrich).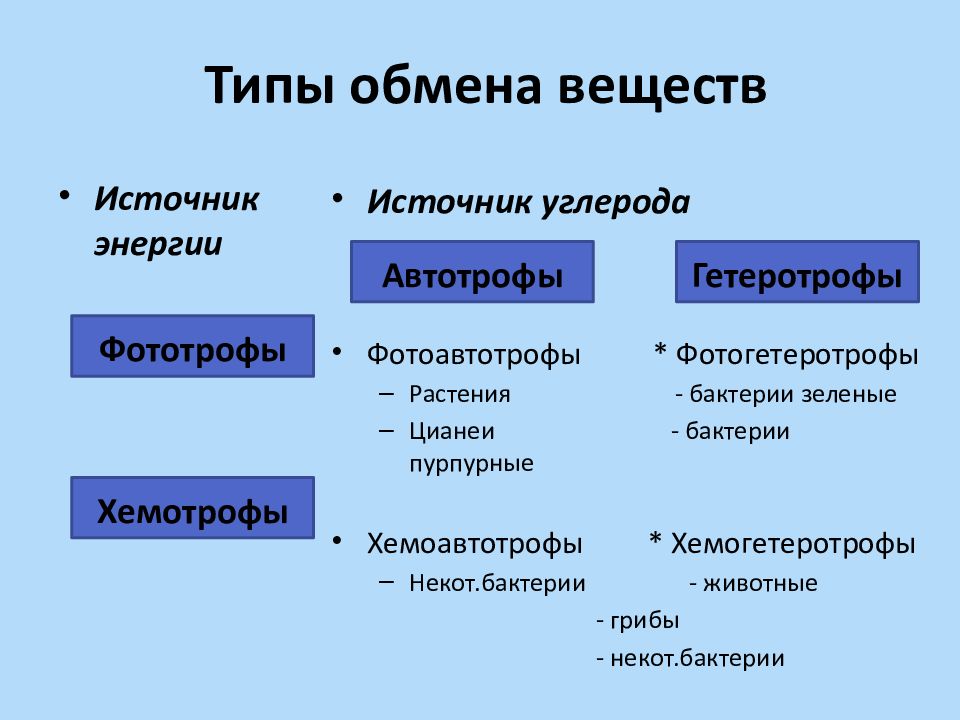 Раствор микроэлементов состоял из 19 г/л Na2ЭДТА⋅2h3O (дигидрат этилендиаминтетраацетата динатрия), 4,5 г/л ZnSO4⋅7h3O, 1 г/л MnCl2⋅4h3O, 0,3 г/л CoCl2⋅6h3O, 0,3 г/л CuSO4⋅5h3O. , 0,4 г/л Na2MoO4⋅2h3O, 4.5 г/л CaCl2⋅2h3O, 3 г/л FeSO4⋅7h3O, 1 г/л h4BO3 и 0,1 г/л йодистого калия. Витаминный раствор состоял из 0,05 г/л d-биотина, 0,2 г/л 4-аминобензойной кислоты, 1 г/л никотиновой кислоты, 1 г/л гемикальциевой соли D-пантотеновой кислоты, 1 г/л пиридоксина-HCl, 1 г/л. L-тиамин-HCl, 25 г/л, и мио-инозитол.
Раствор микроэлементов состоял из 19 г/л Na2ЭДТА⋅2h3O (дигидрат этилендиаминтетраацетата динатрия), 4,5 г/л ZnSO4⋅7h3O, 1 г/л MnCl2⋅4h3O, 0,3 г/л CoCl2⋅6h3O, 0,3 г/л CuSO4⋅5h3O. , 0,4 г/л Na2MoO4⋅2h3O, 4.5 г/л CaCl2⋅2h3O, 3 г/л FeSO4⋅7h3O, 1 г/л h4BO3 и 0,1 г/л йодистого калия. Витаминный раствор состоял из 0,05 г/л d-биотина, 0,2 г/л 4-аминобензойной кислоты, 1 г/л никотиновой кислоты, 1 г/л гемикальциевой соли D-пантотеновой кислоты, 1 г/л пиридоксина-HCl, 1 г/л. L-тиамин-HCl, 25 г/л, и мио-инозитол.
Отбор проб из биореактора.
Для всех моментов времени отбора проб мертвый объем порта отбора проб удалялся перед отбором проб. Определение концентрации сухой массы клеток проводили путем вакуумной фильтрации образцов с использованием предварительно взвешенного 0.Мембрана фильтра 45 мкм (Sartorius Biolabs) с последующей сушкой фильтров в микроволновой печи при мощности 125 Вт в течение 15 мин и хранением фильтров в вакуумном эксикаторе не менее 3 дней. Отбор проб для анализа экзометаболитов проводили путем фильтрации пробы через нейлоновые фильтры с размером пор 0,45 мкм. Бесклеточные образцы хранили при температуре -20 °C до проведения анализа. Для протеомики образцы центрифугировали при <0 °C в течение 5 мин. Осадки клеток промывали фосфатно-солевым буфером, мгновенно замораживали в жидком азоте и хранили при -80°С до анализа.
Бесклеточные образцы хранили при температуре -20 °C до проведения анализа. Для протеомики образцы центрифугировали при <0 °C в течение 5 мин. Осадки клеток промывали фосфатно-солевым буфером, мгновенно замораживали в жидком азоте и хранили при -80°С до анализа.
Статистика и анализ данных.
Анализ всех данных выполнен на основе трех биологических повторностей, за исключением расчетов OUR, CER и RQ, где для S. pombe , S. cerevisiae и RQ использовались данные двух биологических повторностей. К. марксианский .
Анализ экзометаболитов.
Внеклеточная глюкоза, этанол, глицерин, пируват, сукцинат и ацетат были количественно определены с использованием системы высокоэффективной жидкостной хроматографии Ultimate 3000 (Thermo Fisher), оснащенной колонкой BioRad HPX-87H (BioRad) и детектором показателя преломления с 5 мМ H 2 SO 4 в качестве элюирующего буфера при скорости потока 0.6 мл ⋅ мин -1 и температура печи 45 °C.
Анализ газов.
Концентрации кислорода и углекислого газа в выхлопных газах измерялись в режиме реального времени с помощью газоанализатора GA4 (DasGip). OUR в каждый момент времени рассчитывали с поправкой на разбавление образованием углекислого газа, согласно уравнению. 1 :OUR = F*xO2,inVm−xN2,inxN2,out*F*xO2,outVm,[1], где F – расход газа, xO2,in – доля кислорода в воздухе, поступающем в реактор , xO2,вых – доля кислорода в газе, выходящем из реактора, xN2,вх – объединенная доля азота и аргона в воздухе, xN2,вх – объединенная доля азота и аргона, выходящего из реактора, V м — молярный объем при 25 °C (24.465). Общее количество молей кислорода, потребленного до каждой временной точки, рассчитывали как площадь под кривой OUR, а полученные значения наносили на график в зависимости от концентрации биомассы (г сухой массы клеток). Конкретный OUR был рассчитан путем умножения наклона линейной линии тренда на скорость роста. CER рассчитывали по уравнению. 2 :CER = F*xCO2,inVm−F*xCO2,outVm,[2], где xCO2,in и xCO2,out — доля углекислого газа в газе, поступающем в реактор и выходящем из него, соответственно.Дальнейшие расчеты проводились с использованием того же подхода, что и для конкретного OUR.
CER рассчитывали по уравнению. 2 :CER = F*xCO2,inVm−F*xCO2,outVm,[2], где xCO2,in и xCO2,out — доля углекислого газа в газе, поступающем в реактор и выходящем из него, соответственно.Дальнейшие расчеты проводились с использованием того же подхода, что и для конкретного OUR.
Количественные измерения протеома.
Эксперименты по жидкостной хроматографии-МС (ЖХ-МС) проводились на масс-спектрометре Orbitrap Fusion или Orbitrap Fusion Lumos, соединенном с системой Easy-nLC1200 nanoflow LC (все от Thermo Fisher Scientific). Идентификацию пептидов и белков проводили с использованием Proteome Discoverer версии 2.4 (Thermo Fisher Scientific) с Mascot версии 2.5.1 (Matrix Science) в качестве поисковой системы по базе данных.
Количественное определение общего относительного белка между образцами было выполнено с помощью модифицированного метода подготовки образцов с фильтрацией (FASP) (51), который включал двухэтапное расщепление трипсином в буфере 0,5% дезоксихолата натрия/50 мМ бикарбоната триэтиламмония и мечение изобарическими реагентами TMT 10plex (Thermo Fisher Scientific) в соответствии с инструкциями производителя. Опрошенный эталонный образец для каждого вида готовили из аликвот отдельных повторностей и обрабатывали вместе с отдельными образцами.Объединенный набор, меченный ТМТ, был предварительно разделен на 40 первичных фракций с помощью обращенно-фазовой хроматографии с основным рН (bRP-LC) на колонке XBridge BEH C18 (3,5 мкм, 3,0 × 150 мм, Waters Corporation) при рН 10 с использованием Dionex. Система ультравысокоэффективной жидкостной хроматографии Ultimate 3000 (Thermo Fisher Scientific). Наиболее распространенные предшественники пептидов были отобраны в зависимости от данных, спектры диссоциации, индуцированной столкновением MS 2 , для идентификации пептидов были записаны в ионной ловушке, 10 наиболее распространенных ионов-фрагментов были выделены, фрагментированы с использованием диссоциации столкновений с более высокой энергией. , и спектры MS 3 для количественного определения репортерных ионов были записаны в Orbitrap.
Опрошенный эталонный образец для каждого вида готовили из аликвот отдельных повторностей и обрабатывали вместе с отдельными образцами.Объединенный набор, меченный ТМТ, был предварительно разделен на 40 первичных фракций с помощью обращенно-фазовой хроматографии с основным рН (bRP-LC) на колонке XBridge BEH C18 (3,5 мкм, 3,0 × 150 мм, Waters Corporation) при рН 10 с использованием Dionex. Система ультравысокоэффективной жидкостной хроматографии Ultimate 3000 (Thermo Fisher Scientific). Наиболее распространенные предшественники пептидов были отобраны в зависимости от данных, спектры диссоциации, индуцированной столкновением MS 2 , для идентификации пептидов были записаны в ионной ловушке, 10 наиболее распространенных ионов-фрагментов были выделены, фрагментированы с использованием диссоциации столкновений с более высокой энергией. , и спектры MS 3 для количественного определения репортерных ионов были записаны в Orbitrap.
Для количественного определения без использования меток в аликвоту 25 мкг каждого из объединенных эталонных образцов добавляли 5,3 мкг стандарта динамического диапазона протеомики UPS2 (Sigma-Aldrich), расщепленного с использованием протокола FASP, как описано в предыдущем абзаце. и разделяли на восемь фракций с использованием набора спин-колонок с обращенной фазой Pierce с высоким pH (Thermo Fisher Scientific). Каждую пептидную фракцию анализировали в трех повторностях, при этом спектры MS 1 записывали с разрешением 120 000, а спектры MS 2 , зависящие от данных, записывали в ионной ловушке.Данные без меток были обработаны с использованием узла обнаружения признаков Minora в Proteome Discoverer версии 2.4, а значения интенсивности трех повторов инъекции были усреднены. Для оценки абсолютной концентрации белка в объединенном эталонном образце использовали модифицированный подход iBAQ (20). Интенсивность белка делили на количество идентифицированных пептидов, чтобы получить интенсивность iBAQ для каждого белка. Линейная регрессия была рассчитана для взаимосвязи между известным количеством log 10 -трансформированного iBAQ и измеренным количеством log 10 -трансформированного белка в стандартных белках UPS2 с добавлением.
и разделяли на восемь фракций с использованием набора спин-колонок с обращенной фазой Pierce с высоким pH (Thermo Fisher Scientific). Каждую пептидную фракцию анализировали в трех повторностях, при этом спектры MS 1 записывали с разрешением 120 000, а спектры MS 2 , зависящие от данных, записывали в ионной ловушке.Данные без меток были обработаны с использованием узла обнаружения признаков Minora в Proteome Discoverer версии 2.4, а значения интенсивности трех повторов инъекции были усреднены. Для оценки абсолютной концентрации белка в объединенном эталонном образце использовали модифицированный подход iBAQ (20). Интенсивность белка делили на количество идентифицированных пептидов, чтобы получить интенсивность iBAQ для каждого белка. Линейная регрессия была рассчитана для взаимосвязи между известным количеством log 10 -трансформированного iBAQ и измеренным количеством log 10 -трансформированного белка в стандартных белках UPS2 с добавлением. Полученную регрессионную модель и измеренное содержание белков в объединенном эталонном образце с помощью iBAQ использовали для оценки содержания в них белков.
Полученную регрессионную модель и измеренное содержание белков в объединенном эталонном образце с помощью iBAQ использовали для оценки содержания в них белков.
Подробные экспериментальные процедуры, ЖХ-МС и параметры обработки данных описаны в Приложение SI, Дополнительные методы .
Для определения общего содержания белка после протеомного анализа белки экстрагировали кипячением осадка клеток в лизирующем буфере, содержащем 1 М NaOH и 10% додецилсульфата натрия, в течение 10 мин.Собирали супернатант и измеряли концентрацию белка с использованием набора для анализа бихионовой кислоты Pierce Protein Assay Kit (ThermoFisher 23225) и нормализовали по сухой массе клеток.
Ортологический прогноз с помощью OrthoFinder и аннотации GO.
Для идентификации ортологичного белка среди четырех дрожжей, проанализированных в этом исследовании, и для облегчения назначения терминов GO для анализа данных протеомики было проведено сопоставление гомологии всего протеома с использованием OrthoFinder (52). Результаты были обработаны и классифицированы на однокопийные и многокопийные ортологи.Однокопийные ортологи были определены как имеющие один ортолог в каждом из четырех видов, в то время как многокопийные ортологи были определены как имеющие несколько белковых ортологов в одном или нескольких видах. Чтобы присвоить термины GO всем белкам четырех дрожжей, сначала была загружена аннотация терминов GO для S. cerevisiae с использованием пакета biomaRt R (53). Затем эту аннотацию использовали для аннотирования белков S. pombe , K. marxianus и S. stipitis на основе ортологии белков.Аннотации GO использовались для разделения белков на группы по функциям, используемым для анализа на протяжении всего исследования.
Результаты были обработаны и классифицированы на однокопийные и многокопийные ортологи.Однокопийные ортологи были определены как имеющие один ортолог в каждом из четырех видов, в то время как многокопийные ортологи были определены как имеющие несколько белковых ортологов в одном или нескольких видах. Чтобы присвоить термины GO всем белкам четырех дрожжей, сначала была загружена аннотация терминов GO для S. cerevisiae с использованием пакета biomaRt R (53). Затем эту аннотацию использовали для аннотирования белков S. pombe , K. marxianus и S. stipitis на основе ортологии белков.Аннотации GO использовались для разделения белков на группы по функциям, используемым для анализа на протяжении всего исследования.
ФБА.
Модель метаболизма дрожжей ec консенсуса версии 8.3.4 и модели eciSM966 K. marxianus , используемые в этом исследовании, были получены из репозитория GitHub, размещенного в группе (https://github.com/SysBioChalmers/ecModels) (23 ). Модели, специфичные для состояния, были созданы путем включения и ограничения моделей экспериментальными измерениями содержания белка, скорости обмена метаболитов и содержания метаболических ферментов с использованием набора инструментов GECKO (23).Это включало масштабирование коэффициента псевдореакции белка, чтобы он равнялся измеренному содержанию белка, а также масштабирование коэффициента углеводов для поддержания эквивалентного количества массы в псевдореакции биомассы (9). Кроме того, связанное с ростом поддержание, которое в значительной степени отражает затраты на полимеризацию белков и углеводов, было пересчитано, как описано ранее (9). Для моделирования использовался MATLAB R2018b (MathWorks Inc.) с решателем Gurobi (Gurobi Optimizer) в наборе инструментов RAVEN (54).Для моделирования FBA в качестве целевой функции для минимизации была выбрана псевдореакция обмена общего пула белков, и проблема была решена с использованием функцииsolveLP, доступной в наборе инструментов RAVEN.
Модели, специфичные для состояния, были созданы путем включения и ограничения моделей экспериментальными измерениями содержания белка, скорости обмена метаболитов и содержания метаболических ферментов с использованием набора инструментов GECKO (23).Это включало масштабирование коэффициента псевдореакции белка, чтобы он равнялся измеренному содержанию белка, а также масштабирование коэффициента углеводов для поддержания эквивалентного количества массы в псевдореакции биомассы (9). Кроме того, связанное с ростом поддержание, которое в значительной степени отражает затраты на полимеризацию белков и углеводов, было пересчитано, как описано ранее (9). Для моделирования использовался MATLAB R2018b (MathWorks Inc.) с решателем Gurobi (Gurobi Optimizer) в наборе инструментов RAVEN (54).Для моделирования FBA в качестве целевой функции для минимизации была выбрана псевдореакция обмена общего пула белков, и проблема была решена с использованием функцииsolveLP, доступной в наборе инструментов RAVEN. В моделировании ecModel у каждого фермента есть реакция обмена, которая служит показателем потребности в ферменте и имеет единицу ммоль gDW -1 . Использование емкости отдельных ферментов, представляющее долю доступного фермента, которая фактически используется для катализа, рассчитывали как предсказанный in silico обменный поток ферментов, деленный на обилие ферментов, измеренное in vivo.
В моделировании ecModel у каждого фермента есть реакция обмена, которая служит показателем потребности в ферменте и имеет единицу ммоль gDW -1 . Использование емкости отдельных ферментов, представляющее долю доступного фермента, которая фактически используется для катализа, рассчитывали как предсказанный in silico обменный поток ферментов, деленный на обилие ферментов, измеренное in vivo.
Благодарности
Мы благодарим Егора Воронцова из Центра протеомики Сальгренской академии Гетеборгского университета за проведение протеомного анализа. Proteomics Core Facility выражает благодарность Inga-Britt и Arne Lundbergs Forskningsstiftlese за пожертвование прибора Orbitrap Fusion Tribrid MS. Это исследование было поддержано фондом Ново Нордиск и Фондом Кнута и Элис Валленберг. Финансирование открытого доступа предоставляется Технологическим университетом Чалмерса.
Сноски
- Принято 9 ноября 2021 г. проектное исследование; CM, RY и JB провели исследование; К.М., Р.Ю., Дж.Б. и Э.Дж.К. проанализированные данные; СМ. и Р.Ю. написал газету; и Э.Дж.К. и Дж.Н. руководил исследованием.
Авторы заявляют об отсутствии конкурирующих интересов.
Эта статья является прямой отправкой PNAS.
Эта статья содержит вспомогательную информацию в Интернете по адресу https://www.pnas.org/lookup/suppl/doi:10.1073/pnas.2112836118/-/DCSupplemental.
- Copyright © 2021 Автор(ы). Опубликовано ПНАС.
Исключительное обучение, исследования и уход за пациентами
Отделение эндокринологии, метаболизма и питания Университета Герцога является лидером в оказании помощи пациентам с эндокринными заболеваниями.Мы стремимся обучать новое поколение эндокринологов и исследовать причины, лечение и профилактику диабета, эндокринных и метаболических нарушений.
Узнать больше
 проектное исследование; CM, RY и JB провели исследование; К.М., Р.Ю., Дж.Б. и Э.Дж.К. проанализированные данные; СМ. и Р.Ю. написал газету; и Э.Дж.К. и Дж.Н. руководил исследованием.
проектное исследование; CM, RY и JB провели исследование; К.М., Р.Ю., Дж.Б. и Э.Дж.К. проанализированные данные; СМ. и Р.Ю. написал газету; и Э.Дж.К. и Дж.Н. руководил исследованием.Метаболизм во взрослом возрасте не замедляется, как принято считать, исследование показало, что
Винить лишние килограммы в замедлении метаболизма с возрастом? Не так быстро.
Новое международное исследование опровергает распространенное мнение о неизбежном снижении метаболизма во взрослой жизни. Ну, во всяком случае, пока нам не за 60.
Исследователи обнаружили, что пик метаболизма приходится на возраст 1 год, когда дети сжигают калории на 50 процентов быстрее, чем взрослые, а затем постепенно снижается примерно на 3 процента в год примерно до 20 лет. Согласно выводам, опубликованным в четверг в журнале Science.
Согласно выводам, опубликованным в четверг в журнале Science.
Чтобы выявить конкретное влияние возраста на обмен веществ, исследователи скорректировали такие факторы, как размер тела (большие тела сжигают больше калорий в целом, чем маленькие) и обезжиренная мышечная масса (мышцы сжигают больше калорий, чем жир).
«Скорость метаболизма действительно стабильна на протяжении всей взрослой жизни, от 20 до 60 лет», — сказал автор исследования Герман Понцер, адъюнкт-профессор эволюционной антропологии в Университете Дьюка и автор новой книги «Burn» о метаболизме. «Например, мы не видим никакого эффекта менопаузы. И вы знаете, люди скажут: «Ну, когда мне исполнилось 30 лет, мой метаболизм развалился». На самом деле мы не видим никаких доказательств этого».
Понцер и его коллеги изучили базу данных более чем 6400 человек в возрасте от 8 дней до 95 лет из 29 стран мира, которые участвовали в тестах на «воду с двойной маркировкой».С помощью этого метода люди пьют воду, в которой часть водорода и кислорода заменена изотопами этих элементов, которые можно обнаружить в образцах мочи.
«Подсчитав, сколько водорода вы теряете в день и сколько кислорода вы теряете в день, мы можем рассчитать, сколько углекислого газа производит ваше тело каждый день», — объяснил Понцер. «И это очень точное измерение того, сколько калорий вы сжигаете каждый день, потому что вы не можете сжигать калории, не выделяя углекислый газ.
Исследователи проанализировали средние общие ежедневные затраты энергии, которые включают калории, которые мы сжигаем, делая все, от дыхания и переваривания пищи до мышления и движения нашего тела.
«Нет ничего более фундаментального и фундаментального, чем то, как наши тела сжигают энергию, потому что это показывает, как все наши клетки заняты весь день, выполняя свои различные задачи, и у нас не было четкого представления о том, как это меняется в течение дня. всей жизни», — сказал Понцер. «Вам нужны действительно большие наборы данных, чтобы ответить на этот вопрос.И это был первый раз, когда у нас была возможность сделать это с действительно большим набором данных, который позволил бы нам разделить влияние размера тела, возраста, пола и всего прочего на наши расходы энергии в течение дня».
Возьмем, к примеру, тот факт, что скорость обмена веществ у пожилых людей снижается, чего можно было ожидать.
«Люди думали: «Ну, может быть, это потому, что вы менее активны, или, может быть, это потому, что люди склонны терять мышечную массу, когда им за 60, 70 и старше», — сказал он.«Но мы можем исправить все эти вещи. Мы можем сказать: «Нет, нет, нет, это нечто большее». Дело в том, что наши клетки на самом деле меняются».
Результаты не показали резкого увеличения скорости метаболизма в подростковом возрасте или во время беременности, как это принято считать, или что между мужчинами и женщинами существовали определенные различия после учета размера и состава тела.
Какие факторы вызывают увеличение веса?
Зарегистрированный врач-диетолог Коллин Тьюксбери, старший научный сотрудник Пенсильванского университета и представитель Академии питания и диетологии, заявила, что новое исследование вызывает удивление.
«Исторически сложилось так, что при различных изменениях жизненного цикла — полового созревания, беременности, менопаузы — мы думали, что происходит некоторый сдвиг в обмене веществ, и это влияло на статус питания и на то, как мы подходили к вещам с точки зрения питания», — сказала она. . «Эта строгая оценка на высоком уровне этого не показывает».
. «Эта строгая оценка на высоком уровне этого не показывает».
Дело не в том, что прибавка в весе происходит из-за того, что вы больше не «сжигаете те же калории».
Но если изменение метаболизма не играет роли в увеличении веса в определенные моменты взрослой жизни, могут быть и другие способствующие факторы, сказала она.
«Есть много вещей, которые влияют на вес, а также на чье-то питание», — сказал Тьюксбери. «Это не так просто, как один продукт питания или одно изменение образа жизни или одно изменение с биологической точки зрения. Скорее всего, это гораздо более сложная сеть множества различных изменений, происходящих одновременно. Таким образом, это могут быть изменения в рационе питания. Это могут быть изменения в уровне активности. Это может быть место, где они живут, к чему у них есть доступ, каковы изменения их сна».
Стивен Малин, адъюнкт-профессор кинезиологии и здоровья и директор Лаборатории прикладного метаболизма и физиологии Рутгерса, назвал результаты исследования «проливающими свет на то, о чем мы думали, что знаем много, и понимаем, что есть еще что открыть.
Малин сказал, что результаты, например, противоречат убеждению, что у взрослых людей наблюдается снижение метаболизма по мере того, как они переходят от 20 к 30 годам, и что это может способствовать эпидемии ожирения.
«Прибавка в весе происходит не потому, что вы больше не «сжигаете те же калории», — сказал он.
Pontzer сказало заключения в предыдущей жизни выделяет критическую важность младенческого питания соотвествуя увеличивая потребности в энергии растущих младенцев.
In addition, он сказал, результаты изучения смогли иметь значения для сколько людей микстуры в различном времени, когда они smogли быть метаболизируя снадобья по-разному.
В комментарии, опубликованном вместе с новым исследованием, Тимоти Роадс и Розалин Андерсон, работающие в области гериатрии в Университете Висконсина, заявили, что результаты также могут иметь значение для изучения возрастных заболеваний.
«Считается, что снижение с 60 лет отражает изменение тканеспецифического метаболизма, энергии, затрачиваемой на поддержание», — писали они. «Не может быть совпадением, что рост заболеваемости неинфекционными заболеваниями и расстройствами начинается именно в эти же сроки».
«Не может быть совпадением, что рост заболеваемости неинфекционными заболеваниями и расстройствами начинается именно в эти же сроки».
Диабет, ожирение и обмен веществ — Онлайн-библиотека Wiley
Октябрь 2021
Активация или инактивация глюкокиназы: что приведет к лечению диабета 2 типа?
Акинобу Накамура, доктор медицины, Кадзуно Омори, доктор медицины, Ясуо Тераучи, доктор медицины
Сравнимая эффективность и безопасность инсулина гларгина LY2963016 и инсулина гларгина (Lantus®) у китайских пациентов с диабетом 1 типа: фаза III, рандомизированное, контролируемое исследование
Xiang Yan MD, Shan Jiang PhD, Ying Lou PhD, Zhiguang Zhou MD
Dasiglucagon, аналог глюкагона нового поколения, для лечения тяжелой гипогликемии с помощью автоинъектора: результаты фазы 3, рандомизированное, двойное слепое исследование
Timothy S.Бейли, доктор медицины, Джули Уиллард, доктор медицины, Лесли Дж. Клафф, доктор медицины, Дженин Ягер Стоун, MSN, Анита Мелгаард, магистр наук, Рамин Тегеранчи, доктор медицины
Август 2021
Фармакокинетика, безопасность, переносимость и эффективность котадутида, двойного агониста ГПП-1 и рецептора глюкагона, в исследованиях фазы 1/2 у участников азиатского происхождения с избыточным весом или ожирением с СД2 или без него Ким Магистр наук, Роберт А. Гассер-младший, доктор медицинских наук, Даррен Робертсон, доктор философии, Марселла Петроне, доктор философии, Лутц Джермутус, доктор философии, Филип Амбери FRCP
Гассер-младший, доктор медицинских наук, Даррен Робертсон, доктор философии, Марселла Петроне, доктор философии, Лутц Джермутус, доктор философии, Филип Амбери FRCP
Реальные характеристики системы MiniMed™ 670G в Европе
Жюльен Да Сильва, MS, Эмануэле Боси, доктор медицинских наук, Йохан Джендл Проф, Арселия Арриета, MS, Хавьер Кастанеда, MS, Беньямин Гроссман, доктор философии, Тони Л.Кордеро, доктор философии, Джон Шин, доктор философии, Охад Коэн, доктор медицины
СНИЖЕНИЕ РАСПРОСТРАНЕННОСТИ РЕТИНОПАТИИ ПРИ ДИАБЕТЕ 1 ТИПА В ДЕТСТВЕ ЗА ПОСЛЕДНЕЕ ДЕСЯТИЛЕТИЕ , Стефано Цуккини, доктор медицины, Габриэлла Левантини, доктор медицины, Стефано Тумини, доктор медицины, Роберто Франчески, доктор медицины, Виттория Ковин, доктор медицины, Соня Тони, доктор медицины, Елена де Нитто, доктор медицины, Алессандро Сальватони, доктор медицины, Риккардо Скьяффини, доктор медицины
июль 2021
Безопасность и эффективность аналога пептида пролонгированного действия YY при ожирении: рандомизированное плацебо-контролируемое исследование фазы 1
Tricia M. -М. Тан, доктор философии, Джеймс Миннион, доктор философии, Бернард Ху, доктор философии, Лаура-Джейн Болл, бакалавр наук, Решма Мальвия, доктор философии, Эмили Дэй, магистр наук, Франческа Фиорентино, доктор философии, Чарли Бриндли, доктор философии, Джим Буш, доктор философии, Стивен Р. Блум, доктор наук
-М. Тан, доктор философии, Джеймс Миннион, доктор философии, Бернард Ху, доктор философии, Лаура-Джейн Болл, бакалавр наук, Решма Мальвия, доктор философии, Эмили Дэй, магистр наук, Франческа Фиорентино, доктор философии, Чарли Бриндли, доктор философии, Джим Буш, доктор философии, Стивен Р. Блум, доктор наук
Пероральный семаглутид улучшает постпрандиальный метаболизм глюкозы и липидов, а также задерживает опорожнение желудка у пациентов с диабетом 2 типа
Эффективность и безопасность эртуглифлозина у пациентов с сахарным диабетом 2 типа и установленным сердечно-сосудистым заболеванием при использовании инсулина – дополнительное исследование VERTIS CV
июнь 2021
Применение лираглутида 3.0 мг в день при лечении избыточной массы тела и ожирения у людей с шизофренией, шизоаффективным расстройством и первым эпизодом психоза: результаты пилотного рандомизированного двойного слепого плацебо-контролируемого исследования
Clare A. Whicher et al.
Частота и расчетная стоимость консультаций первичной медико-санитарной помощи у людей с диагнозом диабет 2 типа и сопутствующими заболеваниями: ретроспективный анализ 8,9 млн консультаций
Briana Coles et al.
Длительность диабета и гликемический контроль как предикторы сердечно-сосудистых заболеваний и смертности
Fu-Rong Li et al.
Май 2021
Неалкогольная жировая болезнь печени как нарушение обмена веществ у человека: обзор литературы
Bertrand Cariou et al.
HbA1c и здоровье мозга по всему гликемическому спектру
Victoria Garfield et al.
Влияние поливаскулярного заболевания с сопутствующей почечной дисфункцией и без нее на сердечно-сосудистые исходы при диабете: постфактум анализ EMPA-REG OUTCOME
Subodh Verma et al.
НАЖМИТЕ ЗДЕСЬ , чтобы получить доступ ко всем статьям Editor’s Choice!
Лекарственный метаболизм и персонализированная терапия
Главный редактор
Адриан Л. Лерена
Лерена
Центр клинических исследований CICAB, Университетская больница Эстремадура, Бадахос, Испания
allerena@unex.es
Заместитель главного редактора
Вангелис Г. Манолопулос, кафедра фармакологии, Медицинская школа Фракийского университета Демокрита, Александруполис, Греция
Редакционная коллегия
Монжи Бенджеду, кафедра биотехнологии, Университет Западного Кейпа, Кейп Город, Южная Африка
Бинг Чен, Шанхай, кафедра лабораторной медицины, Университет Цзяотун, Шанхай, КНР
Хорхе Дуконге, Медицинский центр, Медицинский кампус Университета Пуэрто-Рико, Сан-Хуан, Пуэрто-Рико
Ярослав Хубачек, Центр экспериментальных исследований Медицина, Институт клинической и экспериментальной медицины, Прага, Чешская Республика
Магнус Ингельман-Сундберг, каф.Физиологии и фармакологии, Каролинский институт, Стокгольм, Швеция
Джорджио Д. Котзалидис, кафедра неврологии, психического здоровья и органов чувств, Университет Ла Сапиенца, Рим, Италия
Яня Марк, фармацевтический факультет, Люблянский университет, Любляна, Словения
Урс А. Мейер, кафедра фармакологии, Биоцентр Базельского университета, Базель, Швейцария
Мейер, кафедра фармакологии, Биоцентр Базельского университета, Базель, Швейцария
Суджит Наир, Фармацевтические науки, Мумбайский университет, Мумбаи, Индия
Чарити Нофцигер, Институт фармакологии и токсикологии, Медицинский университет Парацельса, Зальцбург , Австрия
Ана Пейро, Главный университетский госпиталь Аликанте, Аликанте, Испания
Джорджия Рагия, Лаборатория фармакологии, Фракийский университет Демокрита, Александруполис, Греция
Джае-Гук Шин, Исследовательский центр фармакогеномики, Медицинский колледж Университета Индже, Пусан, Юг Корея
Маурицио Симмако, отд.Неврологии, факультет медицины и психологии, Римский университет Сапиенца, Рим, Италия
Санья Станкович, факультет биологии, Белградский университет, Белград, Сербия
Энрике Теран, Медицинский факультет, Университет Сан-Франциско де Кито, Кито, Эквадор
Рон Х. Н. ван Шайк, кафедра клинической химии, Медицинский центр Эразма, Роттердам, Нидерланды
Софи Висвикис-Сист, INSERM U525, Лотарингский университет, Нанси, Франция
Редакция
Хайке Янке, редактор журнала
Вальтер де Gruyter GmbH
Genthiner Str. 13, 10785 Берлин, Германия
13, 10785 Берлин, Германия
Тел. +49-30-26005-220; Электронная почта: dmpt.editorial@degruyter.com
Отделение эндокринологии и обмена веществ | Департамент медицины Герцога
Отделение эндокринологии, обмена веществ и питания Университета Герцога занимается изучением и лечением наиболее острых проблем, связанных с эндокринной системой, включая диабет, остеопороз, управление липидами, аномалии гипофиза и надпочечников и заболевания щитовидной железы. Читайте нашу последнюю рассылку.
Читайте нашу последнюю рассылку.
Тренируйтесь с нами
Программа стипендийГерцога в области эндокринологии, диабета и обмена веществ предоставляет возможности для повышения квалификации и исследований.
Узнать больше
Узнайте об эндокринологии Duke
Duke входит в число 50 лучших больниц в стране и самую высокопоставленную программу в Северной Каролине по диабету и эндокринологии США.S. News & World Report за 2020–2021 годы.
Подробнее
Последние новости
Новости
Пятница, 14 января 2022 г.
Новости
Четверг, 13 января 2022 г.
Новости
Пятница, 1 октября 2021 г.