Витамин в1 где содержится больше всего: Продукты питания богатые витамином В1
где содержится витамин В1, советы
Витамины – одни из необходимых телу веществ. Без них нельзя представить слаженные, четкие процессы жизнедеятельности. Несмотря на их незаменимость, эти вещества, как правило, не могут быть продуцированы организмом. Витамин В1 также должен ежедневно поставляться вместе с пищей.
Тиамин – водорастворимый витамин, который был открыт в 1910 году.
К сожалению, столь необходимое вещество не может накапливаться в организме, в связи с чем его «запасы» нужно регулярно пополнять
Витамин теряет свойства при температуре свыше 140 градусов и не разрушается в кислотах, что делает его довольно устойчивым.
Назначение вещества
Витамин В1 имеет две основные функции.
- Во-первых, тиамин принимает участие в работе головного мозга и центральной нервной системы. Благодаря ему формируются долговечные нейронные связи, растет способность к запоминанию. Такое свойство делает вещество незаменимым для детей школьного возраста, чья память активно развивается, и пожилых людей, подверженных риску когнитивных нарушений.
 Кроме того, витамин способен облегчить симптомы морской болезни.
Кроме того, витамин способен облегчить симптомы морской болезни. - Во-вторых, без В1 становится невозможным правильное формирование скелета и мышечных тканей. По этой причине его потребление необходимо увеличить спортсменам, стремящимся нарастить массу, людям преклонного возраста, страдающим от болей в костях, и маленьким детям, чей скелет только формируется. Тиамин облегчает зубную боль и положительно влияет на состояние сердца.
 Кроме того, витамин способен облегчить симптомы морской болезни.
Кроме того, витамин способен облегчить симптомы морской болезни.Авитаминоз в данном случае чреват необратимыми последствиями, вплоть до комы и потери памяти. К счастью, до такого доходит редко. Чаще всего дефицит витамина проявляется излишней плаксивостью, раздражительностью, мышечной слабостью.
Продукты, содержащие витамин В1В каких продуктах содержится тиамин
Природа подарила нам огромное разнообразие растительных и животных продуктов, позволяющих получить необходимое количество вещества.
- Свинина. Отличный животный продукт, насыщающий тело большим количеством полезных веществ. Небольшой порции хватит, чтоб восполнить потребность взрослого человека в витамине В1. Из этого мяса можно приготовить множество блюд – достаточно подключить фантазию.
- Горох. Простой и практичный выбор для тех, кто предпочитает растительные продукты. Широко применяется в кулинарии, может служить как гарниром к мясу, так и основой для самостоятельного блюда. Стоит время от времени баловать себя гороховой кашей или супом, чтоб не бояться возможной потери памяти.
- Красная икра. Любители морепродуктов могут порадовать себя красной икрой. Продукт, считающийся деликатесом, скрасит любое застолье, сделав закуски не просто вкусными, но и очень полезными. Стоит помнить, что витамин содержится только в натуральном продукте, а не в искусственных аналогах.
 Небольшой порции хватит, чтоб восполнить потребность взрослого человека в витамине В1. Из этого мяса можно приготовить множество блюд – достаточно подключить фантазию.
Небольшой порции хватит, чтоб восполнить потребность взрослого человека в витамине В1. Из этого мяса можно приготовить множество блюд – достаточно подключить фантазию.- Овсяные отруби. Этот продукт не только полезный, но еще и диетический. Овсяные отруби позволяют контролировать голод, избегать вредных перекусов и насыщают тело необходимым тиамином. Достаточно пару раз в день съедать горстку отрубей, чтобы не знать об авитаминозе.
- Говяжьи почки. Выбор для любителей экспериментов. Несмотря на широкое распространение продукта и его доступность, говяжьи почки не пользуются большой популярностью. И такое отношение совершенно незаслуженно, ведь они очень богаты В1.
 Достаточно пару раз в день съедать горстку отрубей, чтобы не знать об авитаминозе.
Достаточно пару раз в день съедать горстку отрубей, чтобы не знать об авитаминозе.На самом деле, дефицит витамина В1 – довольно редкое явление в современном мире
Это объясняется доступностью пищи и ее разнообразием. Тем не менее, стоит заботиться о своем организме и вовремя обращать внимание на тревожные сигналы.
Еще о витамине В1 и о продуктах, в которых он содержится, в видео:
В каких продуктах содержится витамин B1
Витамин В1 необходим организму человека, поскольку оказывает положительное влияние на функции самых значимых систем организма: нервной, сердечно-сосудистой, эндокринной и пищеварительной. Витамин В1, или тиамин, улучшая циркуляцию крови и помогая обновлению клеток крови, обеспечивает человека жизненной энергией, повышает умственные способности человека, улучшает процессы запоминания, способность к восприятию учебного материала, усиливает иммунитет и защищает организм от негативных внешних воздействий. Итак, зададимся вопросом: в каких продуктах содержится В1?
Витамин В1, или тиамин, улучшая циркуляцию крови и помогая обновлению клеток крови, обеспечивает человека жизненной энергией, повышает умственные способности человека, улучшает процессы запоминания, способность к восприятию учебного материала, усиливает иммунитет и защищает организм от негативных внешних воздействий. Итак, зададимся вопросом: в каких продуктах содержится В1?
Растительные продукты, содержащие B1
Основными растительными продуктами, содержащими витамин B1, являются следующие.
- Овощи: картофель, морковь, брюссельская капуста, брокколи.
- Хлебобулочные изделия, произведенные из муки грубого помола или с добавлением отрубей.
- Бобовые, в частности, фасоль, горох, чечевица.
- Сухофрукты, а именно изюм, чернослив.
- Зелень: шпинат, петрушка.
- Орехи: грецкие, фисташки, арахис.
- Ягоды: голубика, земляника, черная смородина.
- Семечки.
- Крупы: рис, греча, овсянка.
- Отвары трав и плодов: ромашка, мята, плоды шиповника, клевер, крапива
Продукты животного происхождения, имеющие в своем составе B1
Далее рассмотрим, в каких продуктах животного происхождения содержится витамин B1.
- Мясо: свинина, говядина.
- Субпродукты: печень, почки, мозг, сердце.
- Рыба.
- Яйца, в частности, желток.
- Молоко.
Острая нехватка витамина В1 в организме человека, вызванная отсутствием в рационе питания продуктов, содержащих тиамин, может привести к развитию страшной болезни нервной системы под названием бери-бери.
Необходимая суточная норма тиамина для взрослого человека колеблется в интервале от 1,5 до 2,5 мг. В усиленном употреблении продуктов, содержащих тиамин и увеличении суточной дозы витамина В1 нуждаются люди с гиперфункцией щитовидной железы, испытывающие постоянные физические нагрузки, стрессовое воздействие, а также в период болезни и во время выздоровления. В каких случаях необходимо регулярное и достаточное употребление продуктов, которые содержат витамин В1? Прежде всего женщинам в период беременности, детям и пожилым людям.
В заключение хочется отметить, что тиамин является водорастворимым витамином, поэтому легко разрушается. Такие продукты, как чай, кофе, сахар, а также пагубные привычки вроде курения значительно сокращают запасы витамина В1 в организме. Лучше всего происходит усвоение витамина B1 вместе с магнием. Нужно отметить, что магний содержат, например, овсяные и пшеничные отруби, орехи, морские водоросли, креветки.
Такие продукты, как чай, кофе, сахар, а также пагубные привычки вроде курения значительно сокращают запасы витамина В1 в организме. Лучше всего происходит усвоение витамина B1 вместе с магнием. Нужно отметить, что магний содержат, например, овсяные и пшеничные отруби, орехи, морские водоросли, креветки.
Зная, в каких продуктах содержится В1, вы сможете составить сбалансированный рацион питания, укрепить нервную систему, иммунитет и сохранить бодрость и отличное настроение на долгие годы.
Диетолог рассказала о полезных свойствах свинины
По словам врача-диетолога, витамин В1 играет важную роль в углеводном, белковом и жировом обмене. Кроме того, он улучшает умственные способности и настроение, нормализует аппетит, способствует улучшению циркуляции крови, таким образом благотворно влияя на работу сердечно-сосудистой системы.
«В свинине кроме витамина В1 содержатся ненасыщенные жирные кислоты, это линоленовая, линолевая и олеиновая кислоты.
Ранее в Национальном Союзе свиноводов сообщили «ВиЖ», что потребление свинины в России выросло до 77 килограммов на человека в год, это на 5–6% больше, чем в 2019 году. Основная причина – низкая цена по сравнению с говядиной и бараниной.
При этом эксперт обращает внимание, что свинина может отличаться по качеству. На это, по ее словам, влияет целый ряд факторов: возраст свиньи, чем кормили животное, его генотип. «Тенденция последних лет в области селекции – это повышение ценности мяса и увеличение доли постного мяса. То есть снижается жировой компонент и увеличивается белковый. Но одновременно сокращение внутримышечного жира может приводить к ухудшению вкусовых качеств», – указывает специалист.
Кому рекомендуется ограничить потребление свинины
«Употребление в пищу жирных частей этого мяса следует ограничить людям с избыточной массой тела, с заболеваниями поджелудочной железы, с нарушениями липидного обмена и тем, у кого высокий риск развития сердечно-сосудистых заболеваний. Но в целом это не значит, что ее нужно полностью исключить. Нормы потребления свинины у каждого индивидуальны, они зависят от базового метаболизма, физической активности», – пояснила Тамара Прунцева.
В целом диетологи рекомендуют для употребления в пищу свинину, если нет никаких медицинских противопоказаний.
По словам врача, калорийность блюда будет различаться в зависимости от того, отварена ли свинина, приготовлена ли она методом жарки с применением большого количества масла или запечена на гриле. Несмотря на то что диетологи в целом оценивают мясо свинины как продукт, обладающий высокой питательной ценностью, они напоминают, что рацион должен быть сбалансированным.
Правила по выбору свинины
1. Обратить внимание на категорию мяса
Существуют основные категории мяса: категория А и категория Б, которые отличаются между собой по доле мышечной ткани, отмечает диетолог. В категории А доля мышечной ткани составляет более 80%. Продукты с категорией Б содержат массовую долю мышечной ткани от 60 до 80%, они также высокого качества, но, как правило, цена на них чуть ниже.
2. Проверить ветеринарное свидетельство
«Каждый продукт имеет ветеринарное свидетельство, которое вам по запросу могут предоставить в магазине», – пояснила диетолог. Также эксперт рекомендует всегда обращать внимание на срок годности продукта.
3. Оценить цвет мяса
Цвет свинины должен быть от розового до темно-розового. При этом поверхность этого мяса не должна быть липкой. Все это говорит о свежести продукта.
Витамин B1 (тиамин) — Medicine LibreTexts
Тиамин , также известный как тиамин или витамин B 1 , представляет собой витамин, содержащийся в продуктах питания и используемый в качестве пищевой добавки. [2] В качестве добавки используется для лечения и профилактики дефицита тиамина и связанных с ним расстройств, включая бери-бери, корсаковский синдром и корсаковский психоз. Другие применения включают болезнь мочи кленового сиропа и болезнь Ли. Его принимают внутрь или путем инъекций.
[2] В качестве добавки используется для лечения и профилактики дефицита тиамина и связанных с ним расстройств, включая бери-бери, корсаковский синдром и корсаковский психоз. Другие применения включают болезнь мочи кленового сиропа и болезнь Ли. Его принимают внутрь или путем инъекций.
Побочные эффекты, как правило, незначительны. Возможны аллергические реакции, включая анафилаксию. Тиамин входит в группу B-комплексов. Он необходим для метаболизма углеводов. [1] Поскольку люди не могут его вырабатывать, тиамин является важным питательным веществом. Источники питания включают цельнозерновые продукты, мясо и рыбу. [2]
Тиамин был открыт в 1897 г., выделен в 1926 г. и впервые произведен в 1936 г. [3] Он включен в Список основных лекарственных средств Всемирной организации здравоохранения, наиболее эффективных и безопасных лекарственных средств, необходимых в системе здравоохранения. [1] Оптовая стоимость в развивающихся странах составляет около 2,17 долларов США за 1 г флакона. [5] В США месяц замены стоит менее 25 долларов США. [6] В некоторых странах требуется его добавление в определенные продукты, например, в зерновые. [2]
[1] Оптовая стоимость в развивающихся странах составляет около 2,17 долларов США за 1 г флакона. [5] В США месяц замены стоит менее 25 долларов США. [6] В некоторых странах требуется его добавление в определенные продукты, например, в зерновые. [2]
Содержимое
[скрыть]
- 1 Медицинское применение
- 1.1 Дефицит тиамина
- 1.2Другое использование
- 2Побочные эффекты
- 3Химия
- 4Биосинтез
- 5Питание
- 5.1 Наличие в пищевых продуктах
- 5.2 Справочная диетическая норма потребления
- 5.3 Антагонисты
- 6Абсорбция и транспорт
- 6.1Поглощение
- 6.2Связывается с белками сыворотки
- 6.3Клеточное поглощение
- 6.4 Распределение тканей
- 6.5Выделение
- 7Функция
- 7.
- 7.2 Тиамина трифосфат
- 7.3 Аденозинтиаминтрифосфат
- 7.4 Аденозинтиаминдифосфат
- 7.
- 8История
- 9 См. также
- 10Ссылки
- 11Внешние ссылки

Применение в медицине
Дефицит тиамина
См. также: Дефицит тиамина
Тиамин используется для лечения дефицита тиамина, который может привести к летальному исходу. [7] В менее тяжелых случаях неспецифические признаки включают недомогание, потерю веса, раздражительность и спутанность сознания. [8]
Хорошо известные синдромы, вызванные дефицитом тиамина, включают бери-бери, синдром Вернике-Корсакова и невропатию зрительного нерва.
Другое использование
Другие области применения включают болезнь кленового сиропа и болезнь Лея. [1]
Побочные эффекты
Побочные эффекты, как правило, незначительны. [1] Могут возникнуть аллергические реакции, включая анафилаксию. [1]
[1] Могут возникнуть аллергические реакции, включая анафилаксию. [1]
Химия
Тиамин представляет собой бесцветное сероорганическое соединение с химической формулой C 12 H 17 N 4 OS. Его структура состоит из аминопиримидинового и тиазольного колец, связанных метиленовым мостиком. Тиазол замещен метильной и гидроксиэтильной боковыми цепями. Тиамин растворим в воде, метаноле и глицерине и практически не растворим в менее полярных органических растворителях. Он стабилен при кислых pH, но нестабилен в щелочных растворах. [7] [9] Тиамин, представляющий собой N-гетероциклический карбен, может использоваться вместо цианида в качестве катализатора конденсации бензоина. [10] Тиамин нестабилен при нагревании, но стабилен при хранении в замороженном состоянии. [ нужна ссылка ] Он нестабилен при воздействии ультрафиолетового света [9] и гамма-облучения.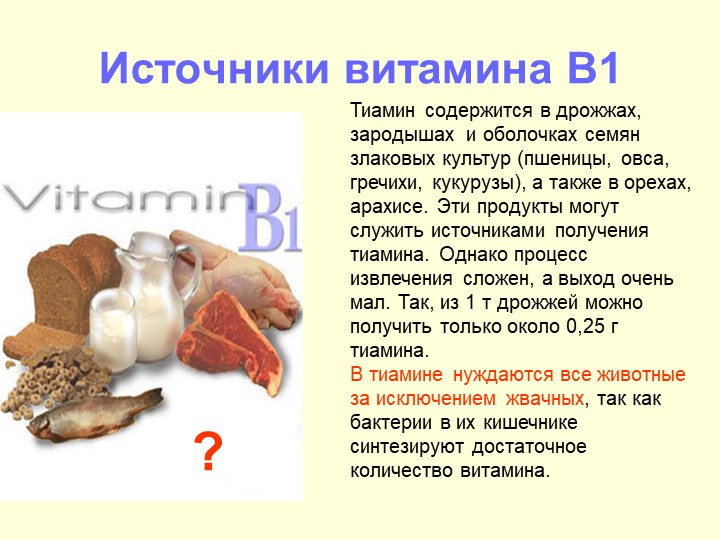 [11] [12] Тиамин сильно реагирует в реакциях Майяра. [7]
[11] [12] Тиамин сильно реагирует в реакциях Майяра. [7]
Биосинтез
Трехмерное изображение рибопереключателя TPP, связанного с тиамином
Сложный биосинтез тиамина происходит в бактериях, некоторых простейших, растениях и грибах. [13] [14] Тиазольный и пиримидиновый фрагменты биосинтезируются отдельно, а затем объединяются с образованием ThMP под действием тиаминфосфатсинтазы (EC 2.5.1.3). Пути биосинтеза могут различаться у разных организмов. У E. coli и других энтеробактерий ThMP может фосфорилироваться до кофактора ThDP с помощью тиаминфосфаткиназы (ThMP + ATP → ThDP + ADP, EC 2.7.4.16). У большинства бактерий и у эукариот ThMP гидролизуется до тиамина, который затем может быть пирофосфорилирован до ThDP с помощью тиаминдифосфокиназы (тиамин + АТФ → ThDP + AMP, EC 2.7.6.2).
Пути биосинтеза регулируются рибопереключателями. Если в клетке присутствует достаточное количество тиамина, то тиамин связывается с мРНК ферментов, которые необходимы для этого пути, и предотвращает их трансляцию. Если тиамина нет, то ингибирования не происходит, и вырабатываются ферменты, необходимые для биосинтеза. Специфический рибопереключатель, рибопереключатель TPP, является единственным рибопереключателем, идентифицированным как у эукариотических, так и у прокариотических организмов. [15]
Если тиамина нет, то ингибирования не происходит, и вырабатываются ферменты, необходимые для биосинтеза. Специфический рибопереключатель, рибопереключатель TPP, является единственным рибопереключателем, идентифицированным как у эукариотических, так и у прокариотических организмов. [15]
Питание
Наличие в пищевых продуктах
Тиамин содержится в большом количестве обработанных и цельных пищевых продуктов, причем съедобные семена, бобовые, рис и обработанные пищевые продукты, такие как сухие завтраки, имеют одно из самых высоких содержаний. [16] [17]
Соль мононитрат тиамина, а не гидрохлорид тиамина, используется для обогащения пищевых продуктов, так как мононитрат более стабилен и не поглощает воду из-за естественной влажности (негигроскопичен), тогда как гидрохлорид тиамина гигроскопичен. [ нужна ссылка ] Когда мононитрат тиамина растворяется в воде, он высвобождает нитрат (около 19% своего веса) и после этого поглощается в виде катиона тиамина.
Некоторые другие продукты, естественно богатые тиамином, — это кукурузная мука, свинина, орехи пекан и шпинат. [16] [17]
Эталонные диетические нормы потребления
Совет по пищевым продуктам и питанию Института медицины США обновил Расчетные средние потребности (EARs) и Рекомендуемые диетические нормы (RDA) для тиамина в 1998 году. Текущие EARs для тиамина для женщин и мужчин в возрасте 14 лет и старше составляют 0,9 мг/день и 1,0 мг/сут соответственно; рекомендуемые суточные дозы составляют 1,1 и 1,2 мг/день. RDA выше, чем EAR, чтобы определить суммы, которые покроют людей с потребностями выше среднего.RDA для беременных составляет 1,4 мг/сутки. RDA для лактации составляет 1,4 мг/день. Для детей до 12 месяцев Адекватное потребление (AI) составляет 0,2-0,3 мг/день. а для детей в возрасте 1–13 лет RDA увеличивается с возрастом с 0,5 до 0,9 мг/день. Что касается безопасности, Совет по пищевым продуктам и питанию Института медицины США устанавливает допустимые верхние уровни потребления (известные как ULs) для витаминов и минералов, когда доказательств достаточно. В случае с тиамином нет UL, так как нет данных о побочных эффектах высоких доз у человека.Европейское управление по безопасности пищевых продуктов рассмотрело тот же вопрос безопасности и также пришло к выводу, что не было достаточных доказательств для установления UL для тиамина. [18] В совокупности EAR, RDA и UL называются эталонными нормами потребления с пищей. [19]
В случае с тиамином нет UL, так как нет данных о побочных эффектах высоких доз у человека.Европейское управление по безопасности пищевых продуктов рассмотрело тот же вопрос безопасности и также пришло к выводу, что не было достаточных доказательств для установления UL для тиамина. [18] В совокупности EAR, RDA и UL называются эталонными нормами потребления с пищей. [19]
Для целей маркировки пищевых продуктов и пищевых добавок в США количество в порции выражается в процентах от дневной нормы (%DV). Для целей маркировки тиамина 100% дневной нормы составляло 1,5 мг, но с мая 2016 года она была изменена на 1.2 мг. Таблица суточной нормы потребления для взрослых до изменения представлена в разделе «Справочная суточная доза». Компании, производящие продукты питания и добавки, должны выполнить это изменение до 28 июля 2018 года.
Антагонисты
Тиамин в пищевых продуктах может расщепляться различными способами. Сульфиты, которые обычно добавляют в пищу в качестве консерванта, [20] атакуют тиамин по метиленовому мостику в структуре, отщепляя пиримидиновое кольцо от тиазольного кольца. [8] Скорость этой реакции увеличивается в кислой среде.Тиамин расщепляется термолабильными тиаминазами (присутствует в сырой рыбе и моллюсках [7] ). Некоторые тиаминазы продуцируются бактериями. Бактериальные тиаминазы представляют собой ферменты клеточной поверхности, которые перед активацией должны диссоциировать от мембраны; диссоциация может происходить у жвачных животных в ацидотических условиях. Бактерии рубца также восстанавливают сульфат до сульфита, поэтому высокое потребление сульфата с пищей может оказывать антагонистическое действие на тиамин.
[8] Скорость этой реакции увеличивается в кислой среде.Тиамин расщепляется термолабильными тиаминазами (присутствует в сырой рыбе и моллюсках [7] ). Некоторые тиаминазы продуцируются бактериями. Бактериальные тиаминазы представляют собой ферменты клеточной поверхности, которые перед активацией должны диссоциировать от мембраны; диссоциация может происходить у жвачных животных в ацидотических условиях. Бактерии рубца также восстанавливают сульфат до сульфита, поэтому высокое потребление сульфата с пищей может оказывать антагонистическое действие на тиамин.
Растительные антагонисты тиамина термостабильны и встречаются как в орто-, так и в пара-гидроксифенолах.Некоторыми примерами этих антагонистов являются кофейная кислота, хлорогеновая кислота и дубильная кислота. Эти соединения взаимодействуют с тиамином, окисляя тиазольное кольцо, что делает его неспособным к абсорбции. Два флавоноида, кверцетин и рутин, также считаются антагонистами тиамина. [8]
Поглощение и транспортировка
Поглощение
Тиамин высвобождается под действием фосфатазы и пирофосфатазы в верхних отделах тонкой кишки. При низких концентрациях процесс опосредуется переносчиком, а при более высоких концентрациях абсорбция происходит посредством пассивной диффузии.Активный транспорт наиболее выражен в тощей и подвздошной кишках; но активный транспорт может подавляться употреблением алкоголя и дефицитом фолиевой кислоты. [7] Снижение всасывания тиамина происходит при приеме выше 5 мг/день. [21] Клетки слизистой оболочки кишечника обладают тиаминпирофосфокиназной активностью, но неясно, связан ли фермент с активным всасыванием. Большая часть тиамина, присутствующего в кишечнике, находится в пирофосфорилированной форме ThDP, но когда тиамин поступает на серозную сторону кишечника, он часто находится в свободной форме.Поглощение тиамина клеткой слизистой оболочки, вероятно, каким-то образом связано с его фосфорилированием/дефосфорилированием. На серозной стороне кишечника данные показали, что выделение витамина этими клетками зависит от Na + -зависимой АТФазы. [8]
При низких концентрациях процесс опосредуется переносчиком, а при более высоких концентрациях абсорбция происходит посредством пассивной диффузии.Активный транспорт наиболее выражен в тощей и подвздошной кишках; но активный транспорт может подавляться употреблением алкоголя и дефицитом фолиевой кислоты. [7] Снижение всасывания тиамина происходит при приеме выше 5 мг/день. [21] Клетки слизистой оболочки кишечника обладают тиаминпирофосфокиназной активностью, но неясно, связан ли фермент с активным всасыванием. Большая часть тиамина, присутствующего в кишечнике, находится в пирофосфорилированной форме ThDP, но когда тиамин поступает на серозную сторону кишечника, он часто находится в свободной форме.Поглощение тиамина клеткой слизистой оболочки, вероятно, каким-то образом связано с его фосфорилированием/дефосфорилированием. На серозной стороне кишечника данные показали, что выделение витамина этими клетками зависит от Na + -зависимой АТФазы. [8]
Связывается с белками сыворотки
Большая часть тиамина в сыворотке связана с белками, главным образом с альбумином. Примерно 90% общего тиамина в крови находится в эритроцитах. Специфический связывающий белок, называемый тиамин-связывающим белком (ТСБ), был идентифицирован в сыворотке крови крыс и считается гормонально-регулируемым белком-носителем, важным для распределения тиамина в тканях. [8]
Примерно 90% общего тиамина в крови находится в эритроцитах. Специфический связывающий белок, называемый тиамин-связывающим белком (ТСБ), был идентифицирован в сыворотке крови крыс и считается гормонально-регулируемым белком-носителем, важным для распределения тиамина в тканях. [8]
Клеточное поглощение
Поглощение тиамина клетками крови и других тканей происходит посредством активного транспорта и пассивной диффузии. [7] Мозгу требуется гораздо больше тиамина, чем другим тканям организма. Большая часть проглоченного тиамина никогда не достигает мозга из-за пассивной диффузии и гематоэнцефалического барьера. Около 80% внутриклеточного тиамина фосфорилировано, и большая его часть связана с белками. В некоторых тканях поглощение и секреция тиамина, по-видимому, опосредованы растворимым переносчиком тиамина, который зависит от Na + и трансцеллюлярного протонного градиента. [8]
Распределение тканей
Запасы тиамина в организме человека составляют от 25 до 30 мг, с наибольшей концентрацией в скелетных мышцах, сердце, головном мозге, печени и почках. ТМФ и свободный (нефосфорилированный) тиамин присутствуют в плазме, молоке, спинномозговой жидкости и, предположительно, во всех внеклеточных жидкостях. В отличие от сильно фосфорилированных форм тиамина, ThMP и свободный тиамин способны пересекать клеточные мембраны. Содержание тиамина в тканях человека меньше, чем у других видов. [8] [22]
ТМФ и свободный (нефосфорилированный) тиамин присутствуют в плазме, молоке, спинномозговой жидкости и, предположительно, во всех внеклеточных жидкостях. В отличие от сильно фосфорилированных форм тиамина, ThMP и свободный тиамин способны пересекать клеточные мембраны. Содержание тиамина в тканях человека меньше, чем у других видов. [8] [22]
Выделение
Тиамин и его кислые метаболиты (2-метил-4-амино-5-пиримидинкарбоновая кислота, 4-метил-тиазол-5-уксусная кислота и тиамин-уксусная кислота) выводятся в основном с мочой. [9]
Функция
Его фосфатные производные участвуют во многих клеточных процессах. Наиболее охарактеризованной формой является тиаминпирофосфат (TPP), кофермент катаболизма сахаров и аминокислот. В дрожжах TPP также требуется на первом этапе спиртового брожения.Все организмы используют тиамин, но он вырабатывается только бактериями, грибами и растениями. Животные должны получать его из своего рациона, и поэтому для людей он является важным питательным веществом. Недостаточное потребление у птиц вызывает характерный полиневрит.
Недостаточное потребление у птиц вызывает характерный полиневрит.
Тиамин обычно считается транспортной формой витамина. Существует пять известных природных производных тиаминфосфата: тиаминмонофосфат (ThMP), тиаминдифосфат (ThDP), также иногда называемый тиаминпирофосфатом (TPP), тиаминтрифосфатом (ThTP) и недавно обнаруженным аденозинтиаминтрифосфатом (AThTP) и аденозинтиамином. дифосфат (АТДФ).В то время как коферментная роль тиаминдифосфата хорошо известна и широко охарактеризована, некоферментное действие тиамина и производных может быть реализовано за счет связывания с рядом недавно идентифицированных белков, которые не используют каталитическое действие тиаминдифосфата [23].
Тиамина дифосфат
Физиологическая роль ThMP неизвестна; однако дифосфат является физиологически значимым. Синтез тиаминдифосфата (ThDP), также известного как тиаминпирофосфат (TPP) или кокарбоксилазы , катализируется ферментом, называемым тиаминдифосфокиназой, в соответствии с реакцией тиамин + АТФ → ThDP + AMP (EC 2. 7.6.2). ThDP является коферментом для нескольких ферментов, которые катализируют перенос двухуглеродных единиц и, в частности, дегидрирование (декарбоксилирование и последующую конъюгацию с коферментом А) 2-оксокислот (альфа-кетокислоты). Примеры включают:
7.6.2). ThDP является коферментом для нескольких ферментов, которые катализируют перенос двухуглеродных единиц и, в частности, дегидрирование (декарбоксилирование и последующую конъюгацию с коферментом А) 2-оксокислот (альфа-кетокислоты). Примеры включают:
- Присутствует у большинства видов
- пируватдегидрогеназа и 2-оксоглутаратдегидрогеназа (также называемая α-кетоглутаратдегидрогеназой)
- дегидрогеназа α-кетокислот с разветвленной цепью
- 2-гидроксифитаноил-КоА-лиаза
- транскетолаза
- Присутствует у некоторых видов:
- пируватдекарбоксилаза (в дрожжах)
- несколько дополнительных бактериальных ферментов
Ферменты транскетолаза, пируватдегидрогеназа (ПДГ) и 2-оксоглутаратдегидрогеназа (ОГДГ) играют важную роль в углеводном обмене.Цитозольный фермент транскетолаза играет ключевую роль в пентозофосфатном пути, основном пути биосинтеза пентозных сахаров дезоксирибозы и рибозы. Митохондриальные PDH и OGDH являются частью биохимических путей, которые приводят к образованию аденозинтрифосфата (АТФ), который является основной формой энергии для клетки. PDH связывает гликолиз с циклом лимонной кислоты, в то время как реакция, катализируемая OGDH, является лимитирующей стадией цикла лимонной кислоты. В нервной системе ПДГ также участвует в производстве ацетилхолина, нейротрансмиттера, и в синтезе миелина. [24]
Митохондриальные PDH и OGDH являются частью биохимических путей, которые приводят к образованию аденозинтрифосфата (АТФ), который является основной формой энергии для клетки. PDH связывает гликолиз с циклом лимонной кислоты, в то время как реакция, катализируемая OGDH, является лимитирующей стадией цикла лимонной кислоты. В нервной системе ПДГ также участвует в производстве ацетилхолина, нейротрансмиттера, и в синтезе миелина. [24]
Тиамина трифосфат
Трифосфат тиамина (ThTP) долгое время считался особой нейроактивной формой тиамина. Однако недавно было показано, что ThTP существует в бактериях, грибах, растениях и животных, что предполагает гораздо более общую клеточную роль. [25] В частности, в E. coli он, по-видимому, играет роль в ответ на дефицит аминокислот. [26]
Аденозинтиаминтрифосфат
Аденозинтиаминтрифосфат (АТТФ) или тиаминилированный аденозинтрифосфат недавно был обнаружен в Escherichia coli , где он накапливается в результате углеродного голодания. [27] В E. coli AThTP может составлять до 20% общего тиамина. Он также содержится в меньших количествах в дрожжах, корнях высших растений и тканях животных. [28]
[27] В E. coli AThTP может составлять до 20% общего тиамина. Он также содержится в меньших количествах в дрожжах, корнях высших растений и тканях животных. [28]
Аденозинтиаминдифосфат
Аденозинтиаминдифосфат (АТДФ) или тиаминилированный аденозиндифосфат присутствует в небольших количествах в печени позвоночных, но его роль остается неизвестной. [28]
История
Тиамин был первым из описанных водорастворимых витаминов, [7] , что привело к открытию большего количества таких микроэлементов, необходимых для выживания, и к понятию витаминов.
В 1884 году Канехиро Такаки (1849–1920), главный хирург японского флота, отверг предыдущую теорию микробов бери-бери и вместо этого предположил, что болезнь возникает из-за недостаточного питания. [29] Сменив диету на военном корабле, он обнаружил, что замена диеты, состоящей только из белого риса, на диету, содержащую также ячмень, мясо, молоко, хлеб и овощи, почти устранила авитаминоз во время 9-месячного морского путешествия. Тем не менее, Такаки добавил много продуктов в успешную диету и неправильно приписал пользу повышенному потреблению азота, поскольку в то время витамины были неизвестными веществами.Военно-морской флот также не был убежден в необходимости такой дорогостоящей программы улучшения питания, и многие мужчины продолжали умирать от авитаминоза даже во время русско-японской войны 1904–1905 годов. Лишь в 1905 году, после того как в рисовых отрубях (удаленных путем полировки до белого риса) и в коричневом ячменном рисе был обнаружен фактор против бери-бери, эксперимент Такаки был вознагражден, сделав его бароном в японской системе пэров, после чего он был нежно по прозвищу «Ячменный барон».
Тем не менее, Такаки добавил много продуктов в успешную диету и неправильно приписал пользу повышенному потреблению азота, поскольку в то время витамины были неизвестными веществами.Военно-морской флот также не был убежден в необходимости такой дорогостоящей программы улучшения питания, и многие мужчины продолжали умирать от авитаминоза даже во время русско-японской войны 1904–1905 годов. Лишь в 1905 году, после того как в рисовых отрубях (удаленных путем полировки до белого риса) и в коричневом ячменном рисе был обнаружен фактор против бери-бери, эксперимент Такаки был вознагражден, сделав его бароном в японской системе пэров, после чего он был нежно по прозвищу «Ячменный барон».
Конкретная связь с зерном была установлена в 1897 году Христианом Эйкманом (1858–1930), военным врачом в Голландской Индии, который обнаружил, что у птиц, питавшихся вареным полированным рисом, развился паралич, который можно было вылечить, прекратив полировку риса. . [30] Он приписал бери-бери нервному яду в эндосперме риса, от которого наружные слои зерна давали защиту организму. Его коллега Геррит Грийнс (1865–1944) правильно интерпретировал связь между чрезмерным потреблением шлифованного риса и авитаминозом в 1901 году: он пришел к выводу, что рис содержит важные питательные вещества во внешних слоях зерна, которые удаляются при шлифовке. [31]
Его коллега Геррит Грийнс (1865–1944) правильно интерпретировал связь между чрезмерным потреблением шлифованного риса и авитаминозом в 1901 году: он пришел к выводу, что рис содержит важные питательные вещества во внешних слоях зерна, которые удаляются при шлифовке. [31]
В конце концов Эйкману была присуждена Нобелевская премия по физиологии и медицине в 1929 году за то, что его наблюдения привели к открытию витаминов.Эти соединения были названы польским биохимиком Казимиром Функом. В 1911 году Казимир Функ выделил противоневритическое вещество из рисовых отрубей, которое он назвал «витамином» (из-за того, что оно содержит аминогруппу). Голландские химики Баренд Коенрад Петрус Янсен (1884–1962) и его ближайший сотрудник Виллем Фредерик Донат (1889–1957) в 1926 году выделили и кристаллизовали активное вещество [32] , структура которого была определена Робертом Раннелсом Уильямсом. (1886–1965), американский химик, в 1934 г.Тиамин («серосодержащий витамин») был синтезирован в 1936 г. той же группой. [33]
той же группой. [33]
Тиамин был впервые назван «аневрином» (антиневритическим витамином). [34] Сэр Рудольф Питерс из Оксфорда представил голубей, лишенных тиамина, в качестве модели для понимания того, как дефицит тиамина может привести к патологическим и физиологическим симптомам бери-бери. Действительно, кормление голубей полированным рисом приводит к легко распознаваемому поведению втягивания головы, состоянию, называемому опистотонусом.Если не лечить, животное погибнет через несколько дней. Введение тиамина на стадии опитотонуса приводит к полному излечению животного в течение 30 мин. Поскольку в мозге голубей до и после обработки тиамином не наблюдалось морфологических изменений, Петерс ввел понятие биохимического поражения. [35]
Когда Lohman и Schuster (1937) показали, что дифосфорилированное производное тиамина (тиаминдифосфат, ThDP) является кофактором, необходимым для окислительного декарбоксилирования пирувата, [36] (реакция, которая, как теперь известно, катализируется пируватдегидрогеназой), механизм действия тиамина в клеточном метаболизме, казалось бы, выяснен. В настоящее время эта точка зрения кажется чрезмерно упрощенной: пируватдегидрогеназа является лишь одним из нескольких ферментов, требующих тиаминдифосфата в качестве кофактора; кроме того, с тех пор были обнаружены другие производные тиаминфосфата, и они также могут способствовать симптомам, наблюдаемым при дефиците тиамина.
В настоящее время эта точка зрения кажется чрезмерно упрощенной: пируватдегидрогеназа является лишь одним из нескольких ферментов, требующих тиаминдифосфата в качестве кофактора; кроме того, с тех пор были обнаружены другие производные тиаминфосфата, и они также могут способствовать симптомам, наблюдаемым при дефиците тиамина.
Наконец, Рональд Бреслоу в 1958 году объяснил механизм, с помощью которого тиаминовая часть ThDP проявляет свою коферментную функцию путем замещения протона в положении 2 тиазолиевого кольца. [37]
дозировка витамина B1 (тиамина), показания, взаимодействие, побочные эффекты и многое другое
Минорный (1) амикацин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тиамин усиливает эффекты атракурия за счет неустановленного механизма взаимодействия. Незначительное/значение неизвестно.
Незначительное/значение неизвестно.
Тщательный мониторинг (1) азитромицин снижает уровень или эффект тиамина, изменяя кишечную флору.Относится только к пероральной форме обоих средств. Используйте осторожность/монитор.
Незначительный (1)азтреонам снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)буметанид снижает уровень тиамина за счет увеличения почечного клиренса. Незначительное/значение неизвестно.
Минорный (1) кофеин снижает уровень тиамина за счет ингибирования всасывания в желудочно-кишечном тракте.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно. Кофе, чай богаты антитиаминовыми факторами.
Минорный (1) цефадроксил снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) цефамандол снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) цефпиром снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1) цефтибутен снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)цефалексин снижает уровень или эффект тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Незначительное (1)хлоргексидин перорально снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Незначительная (1) ципрофлоксацин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тиамин усиливает действие цисатракурия за счет неустановленного механизма взаимодействия.Незначительное/значение неизвестно.
Тщательный мониторинг (1) кларитромицин снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств. Используйте осторожность/монитор.
Незначительная (1) клиндамицин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) дапсон снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) демеклоциклин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) доксициклин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1)эртапенем снижает уровень или действие тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Тщательный мониторинг (1) основание эритромицина снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств. Используйте осторожность/монитор.
Тщательный мониторинг (1) этилсукцинат эритромицина снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств.Используйте осторожность/монитор.
Тщательный мониторинг (1) эритромицина лактобионат снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств. Используйте осторожность/монитор.
Тщательный мониторинг (1) стеарат эритромицина снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств. Используйте осторожность/монитор.
Минорная (1)этакриновая кислота снижает уровень тиамина за счет увеличения почечного клиренса.Незначительное/значение неизвестно.
Минорный (1)этанол снижает уровень тиамина за счет ингибирования всасывания в желудочно-кишечном тракте. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно. Добавка тиамина рекомендуется при алкоголизме.
Незначительная (1)флероксацин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Незначительный (1)фосфомицин снижает уровень или эффект тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1) фуросемид снижает уровень тиамина за счет увеличения почечного клиренса. Незначительное/значение неизвестно.
Малый (1) гемифлоксацин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) гентамицин снижает уровень или эффект тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Незначительная (1)левофлоксацин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)линезолид снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)меропенем снижает уровень или действие тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1)меропенем/ваборбактам снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)метронидазол снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1) миноциклин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Незначительная (1)моксифлоксацин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)нафциллин снижает уровень или эффект тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Незначительное (1)неомицин ПО снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)нитрофурантоин снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1) офлоксацин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Тщательный мониторинг (1) тиамин снижает уровень или эффект омадациклина за счет ингибирования всасывания в желудочно-кишечном тракте. Относится только к пероральной форме обоих средств. Изменить терапию/внимательно контролировать. Продукты, содержащие поливалентные катионы, могут нарушать всасывание тетрациклинов, что может снижать их эффективность. Раздельное дозирование тетрациклинов от этих продуктов.
Минорный (1)тиамин усиливает эффекты onabotulinumtoxinA за счет неустановленного механизма взаимодействия.Незначительное/значение неизвестно.
Минорный (1)окситетрациклин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тиамин усиливает действие панкурония за счет неуказанного механизма взаимодействия. Незначительное/значение неизвестно.
Незначительное/значение неизвестно.
Небольшой (1)паромомицин снижает уровень или действие тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1)пивмециллинам снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) хинупристин/дальфопристин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тиамин усиливает действие рапакурония за счет неустановленного механизма взаимодействия. Незначительное/значение неизвестно.
Минорный (1)тиамин усиливает действие рокурония за счет неустановленного механизма взаимодействия. Незначительное/значение неизвестно.
Незначительное/значение неизвестно.
Тщательный мониторинг (1) рокситромицин снижает уровень или эффект тиамина, изменяя кишечную флору. Относится только к пероральной форме обоих средств.Используйте осторожность/монитор.
Малый (1) стрептомицин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тиамин усиливает действие сукцинилхолина за счет неуказанного механизма взаимодействия. Незначительное/значение неизвестно.
Малый (1) сульфадиазин снижает уровень или эффект тиамина, изменяя кишечную флору.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) сульфаметоксазол снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1) сульфизоксазол снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1) тейкопланин снижает уровень или эффект тиамина за счет изменения кишечной флоры.Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1)темоциллин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тетрациклин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тикарциллин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1)тигециклин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Малый (1) тобрамицин снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)торсемид снижает уровень тиамина за счет увеличения почечного клиренса.Незначительное/значение неизвестно.
Минорный (1)триметоприм снижает уровень или действие тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Второстепенный (1) ванкомицин снижает уровень или эффект тиамина за счет изменения кишечной флоры. Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Относится только к пероральной форме обоих средств. Незначительное/значение неизвестно.
Минорный (1)тиамин усиливает действие векурония за счет неуказанного механизма взаимодействия.Незначительное/значение неизвестно.
Витамин B1 (тиамин) — Nuts.com
Что такое тиамин (витамин B1)?
Тиамин — это витамин, также известный как витамин B1 или тиамин. Он был назван витамином B1, потому что это был первый витамин B, обнаруженный в организме. Он наиболее известен как тиамин, хотя его также можно назвать тиамином, гидрохлоридом тиамина или мононитратом тиамина. Тиамин можно найти как в растениях, так и в животных, и, как и все витамины группы В, он водорастворим, то есть не накапливается в организме.
Преимущества витамина B1
Тиамин используется для многих различных пищевых преимуществ. Он играет решающую роль в некоторых метаболических реакциях. Он необходим для образования аденозинтрифосфата (АТФ), который используется для получения энергии каждой клеткой тела. Эта энергия берется из пищевых углеводов и жиров. Витамин B1 также известен как «антистрессовый» витамин, потому что часто считается, что он помогает укрепить иммунную систему и повысить устойчивость организма к стрессовым ситуациям.Тиамин также можно использовать при проблемах с пищеварением, включая неправильное питание, диарею и язвенный колит. Кроме того, витамин B1 использовался по целому ряду медицинских причин, таких как СПИД, боли при диабете, болезни сердца, старение, повреждение головного мозга, укачивание и некоторые проблемы со зрением. Люди также использовали тиамин для поддержания позитивного настроя, улучшения обучения, увеличения энергии, снижения стресса и улучшения спортивных результатов.
Эта энергия берется из пищевых углеводов и жиров. Витамин B1 также известен как «антистрессовый» витамин, потому что часто считается, что он помогает укрепить иммунную систему и повысить устойчивость организма к стрессовым ситуациям.Тиамин также можно использовать при проблемах с пищеварением, включая неправильное питание, диарею и язвенный колит. Кроме того, витамин B1 использовался по целому ряду медицинских причин, таких как СПИД, боли при диабете, болезни сердца, старение, повреждение головного мозга, укачивание и некоторые проблемы со зрением. Люди также использовали тиамин для поддержания позитивного настроя, улучшения обучения, увеличения энергии, снижения стресса и улучшения спортивных результатов.
Диетические источники
Большинство пищевых продуктов содержат следовые количества тиамина.Тем не менее, более крупные источники витамина B1 включают свинину, говядину, птицу и мясные субпродукты. Другими богатыми диетическими источниками являются цельнозерновые злаки и рис, орехи, бобовые, отруби, дрожжи и зародыши пшеницы. Вы также можете принимать тиамин в качестве добавки. Обычно он содержится в поливитаминах и комплексах витаминов группы В. Этот витаминный комплекс обычно включает витамин В1, а также витамин В2 (рибофлавин), витамин В3 (ниацин), витамин В5 (пантотеновая кислота), витамин В6 (пиридоксин) и витамин В12 (цианокобаламин). Эти добавки выпускаются в различных формах, включая жевательные таблетки, мягкие гели, пастилки и в жидкой форме.Обязательно проверьте этикетку, так как витамин B1 также может быть написан как гидрохлорид тиамина или мононитрат тиамина.
Вы также можете принимать тиамин в качестве добавки. Обычно он содержится в поливитаминах и комплексах витаминов группы В. Этот витаминный комплекс обычно включает витамин В1, а также витамин В2 (рибофлавин), витамин В3 (ниацин), витамин В5 (пантотеновая кислота), витамин В6 (пиридоксин) и витамин В12 (цианокобаламин). Эти добавки выпускаются в различных формах, включая жевательные таблетки, мягкие гели, пастилки и в жидкой форме.Обязательно проверьте этикетку, так как витамин B1 также может быть написан как гидрохлорид тиамина или мононитрат тиамина.
Рекомендуемая суточная доза
Поскольку привычки в питании и образе жизни могут повлиять на необходимое ежедневное потребление витаминов, важно проконсультироваться с врачом, чтобы узнать, какое количество лучше всего подходит для вас. В качестве руководства Национальная академия наук рекомендует следующую суточную дозу тиамина:
- Младенцы:
- Новорожденные, 6 месяцев: 0.2 мг
- Младенцы от 7 месяцев до 1 года: 0,3 мг
- Дети от 1 до 3 лет: 0,5 м
- Маленькие дети:
- Дети от 4 до 8 лет: 0,6 мг
- Дети от 9 до 13 лет: 0,9 мг
- Подростки:
- Мужчины от 14 до 18 лет: 1,2 мг
- Женщины от 14 до 18 лет: 1 мг
- Взрослые:
- Мужчины от 19 лет и старше: 1. 2 мг
- Женщины 19 лет и старше: 1,1 мг
- Беременные или кормящие женщины: 1,4 мг
- Мужчины от 19 лет и старше: 1.
 2 мг
2 мгДефицит тиамина
Поскольку тиамин присутствует во многих продуктах, большинство людей получают достаточное количество витамина B1 в своем рационе. Тем не менее, есть некоторые факторы, которые увеличивают риск дефицита. Люди, страдающие болезнью Крона или анорексией, подвержены большему риску нехватки витамина B1.Алкоголики или лица, проходящие диализ почек, также могут иметь дефицит. Кроме того, увеличение потребления обработанных пищевых продуктов также может вызвать повышенный риск низкого уровня тиамина. Витамин B1 является одним из питательных веществ, который чаще всего удаляется из пищи при современной пищевой промышленности. Симптомы дефицита тиамина могут включать головную боль, тошноту, усталость, депрессию и боль в животе. Хотя в Соединенных Штатах это редкость, важно знать о дефиците витамина B1. Известно, что тяжелые и длительные случаи дефицита витамина B1 влияют на сердце, нервную систему и пищеварительную функцию.
| тиамин | витамин В 1 | компонент кофермента углеводного обмена; поддерживает нормальную нервную функцию | поражение нервов и истощение сердечной мышцы |
| рибофлавин | витамин В 2 | компонент коферментов, необходимых для производства энергии и метаболизма липидов, витаминов, минералов и лекарств; антиоксидант | воспаление кожи, языка и губ; нарушения зрения; нервные симптомы |
| ниацин | никотиновая кислота, никотинамид | компонент коферментов, широко используемых в клеточном метаболизме, окислении топливных молекул и синтезе жирных кислот и стероидов | поражения кожи, желудочно-кишечные расстройства, нервные симптомы |
| витамин В 6 | пиридоксин, пиридоксаль, пиридоксамин | компонент коферментов метаболизма аминокислот и других азотсодержащих соединений; синтез гемоглобина, нейромедиаторов; регуляция уровня глюкозы в крови | дерматит, психическая депрессия, спутанность сознания, судороги, анемия |
| фолиевая кислота | фолат, фолацин, птероилглутаминовая кислота | компонент коферментов в синтезе ДНК, метаболизме аминокислот; необходим для деления клеток, созревания эритроцитов | нарушение образования эритроцитов, слабость, раздражительность, головная боль, сердцебиение, воспаление ротовой полости, дефекты нервной трубки у плода |
| витамин В 12 | кобаламин, цианокобаламин | кофактор ферментов метаболизма аминокислот (включая фолиевую кислоту) и жирных кислот; необходим для синтеза новых клеток, нормального кроветворения и неврологической функции | гладкость языка, желудочно-кишечные расстройства, нервные симптомы |
| пантотеновая кислота | как компонент кофермента А, необходимого для метаболизма углеводов, белков и жиров; кофактор удлинения жирных кислот | слабость, желудочно-кишечные расстройства, нервные симптомы, повышенная утомляемость, нарушения сна, беспокойство, тошнота | |
| биотин | кофактор метаболизма углеводов, жирных кислот и аминокислот | дерматит, выпадение волос, конъюнктивит, неврологические симптомы | |
| Витамин С | аскорбиновая кислота | антиоксидант; синтез коллагена, карнитина, аминокислот и гормонов; иммунная функция; усиливает усвоение негемового железа (из растительной пищи) | припухлость и кровоточивость десен, болезненность и скованность суставов и нижних конечностей, кровоизлияния под кожу и в глубокие ткани, медленное заживление ран, анемия |
| витамин А | ретинол, ретиналь, ретиноевая кислота, бета-каротин (растительная версия) | нормальное зрение, целостность эпителиальных клеток (слизистых оболочек и кожи), размножение, эмбриональное развитие, рост, иммунный ответ | нарушения зрения, приводящие к слепоте, задержке роста, сухости кожи, диарее, восприимчивости к инфекциям |
| Витамин Д | кальциферол, калатриол (1,25-дигидроксивитамин D 1 или гормон витамина D), холекальциферол (D 3 ; растительная версия), эргокальциферол (D 2 ; животная версия) | поддержание уровня кальция и фосфора в крови, правильная минерализация костей | нарушение роста костей у детей, мягкие кости у взрослых |
| витамин Е | альфа-токоферол, токоферол, токотриенол | антиоксидант; прерывание свободнорадикальных цепных реакций; защита полиненасыщенных жирных кислот, клеточных мембран | периферическая невропатия, разрушение эритроцитов |
| витамин К | филлохинон, менахинон, менадион, нафтохинон | синтез белков, участвующих в процессах свертывания крови и костного метаболизма | нарушение свертываемости крови и внутреннее кровотечение |
Границы | Изучение круговорота витамина B1 и его связей с микробным сообществом в северной части Атлантического океана
Введение
Витамин B1, также известный как тиамин, является важным коферментом, необходимым для метаболизма углерода во всех сферах жизни (Monteverde et al. , 2017). Наличие витамина B1 может контролировать активность морских микробов, взаимодействие и структуру сообщества. В морских системах в микробных геномах обнаружено более 30 зависимых от витамина В1 ферментов (Schowen, 1998; Sañudo-Wilhelmy et al., 2014). Эти ферменты в основном используются для метаболизма углеводов и аминокислот с разветвленной цепью и включают пируватдегидрогеназу, транскетолазу и оксоглутаратдегидрогеназу, которые катализируют ключевые стадии циклов ТСА и Кальвина-Бенсона (Rapala-Kozik, 2011).Этот кофермент действует путем стабилизации ацилкарбаниона в реакциях перегруппировки (Jurgenson et al., 2009). Витамин B1 имеет гетероциклическую структуру, состоящую из пиримидинового и тиазольного кольца (Begley et al., 1999; Chatterjee et al., 2006; Jurgenson et al., 2009). Эти кольца биосинтезируются отдельными метаболическими путями, а затем лигируются вместе с образованием тиамина, витамина B1 (Jurgenson et al., 2009). Пути этого процесса биосинтеза различаются у бактерий (Begley et al.
, 2017). Наличие витамина B1 может контролировать активность морских микробов, взаимодействие и структуру сообщества. В морских системах в микробных геномах обнаружено более 30 зависимых от витамина В1 ферментов (Schowen, 1998; Sañudo-Wilhelmy et al., 2014). Эти ферменты в основном используются для метаболизма углеводов и аминокислот с разветвленной цепью и включают пируватдегидрогеназу, транскетолазу и оксоглутаратдегидрогеназу, которые катализируют ключевые стадии циклов ТСА и Кальвина-Бенсона (Rapala-Kozik, 2011).Этот кофермент действует путем стабилизации ацилкарбаниона в реакциях перегруппировки (Jurgenson et al., 2009). Витамин B1 имеет гетероциклическую структуру, состоящую из пиримидинового и тиазольного кольца (Begley et al., 1999; Chatterjee et al., 2006; Jurgenson et al., 2009). Эти кольца биосинтезируются отдельными метаболическими путями, а затем лигируются вместе с образованием тиамина, витамина B1 (Jurgenson et al., 2009). Пути этого процесса биосинтеза различаются у бактерий (Begley et al. , 1999), археи (Maupin-Furlow, 2018) и эукариоты (Chatterjee et al., 2006). Биосинтез витамина B1 регулируется рибопереключателями, которые считаются древними регуляторными структурами, которые делают биосинтез B1 чувствительным к внутриклеточным концентрациям различных форм витамина B1, его предшественников и продуктов распада (Winkler and Breaker, 2005; Croft et al., 2007). ; McRose et al., 2014; Atilho et al., 2019). Транспортные белки клеточной мембраны для витамина B1 распространены у морских микробов, что указывает на то, что некоторые члены микробного сообщества могут ассимилировать экзогенный витамин B1 и родственные ему соединения из растворенного пула (Gómez-Consarnau et al., 2016; Донован и др., 2018).
, 1999), археи (Maupin-Furlow, 2018) и эукариоты (Chatterjee et al., 2006). Биосинтез витамина B1 регулируется рибопереключателями, которые считаются древними регуляторными структурами, которые делают биосинтез B1 чувствительным к внутриклеточным концентрациям различных форм витамина B1, его предшественников и продуктов распада (Winkler and Breaker, 2005; Croft et al., 2007). ; McRose et al., 2014; Atilho et al., 2019). Транспортные белки клеточной мембраны для витамина B1 распространены у морских микробов, что указывает на то, что некоторые члены микробного сообщества могут ассимилировать экзогенный витамин B1 и родственные ему соединения из растворенного пула (Gómez-Consarnau et al., 2016; Донован и др., 2018).
Несмотря на универсальную метаболическую потребность морских микроорганизмов в витамине B1, у многих организмов отсутствует полный путь биосинтеза витамина B1, и поэтому они являются ауксотрофами витамина B1. Ауксотрофность витамина В1 в фитопланктоне была обнаружена в ходе ранних работ по культивированию (Hunter and Provasoli, 1964; Provasoli and Carlucci, 1974). Карлуччи и др. эти результаты были подтверждены с помощью метода биоанализа для определения концентрации витамина В1 как в растворенном виде в морской воде, так и внутри клеток фитопланктона (Carlucci, 1970; Carlucci and Bowes, 1972).Секвенирование генома оказалось особенно эффективным для выявления ауксотрофии витамина B1 в морских системах. На основе геномов и коллекций клеточных культур было подсчитано, что около 20% бактериопланктона и 30–50% фитопланктона обязательно нуждаются в экзогенных источниках витамина B1 (Tang et al., 2010; Sañudo-Wilhelmy et al., 2014).
Карлуччи и др. эти результаты были подтверждены с помощью метода биоанализа для определения концентрации витамина В1 как в растворенном виде в морской воде, так и внутри клеток фитопланктона (Carlucci, 1970; Carlucci and Bowes, 1972).Секвенирование генома оказалось особенно эффективным для выявления ауксотрофии витамина B1 в морских системах. На основе геномов и коллекций клеточных культур было подсчитано, что около 20% бактериопланктона и 30–50% фитопланктона обязательно нуждаются в экзогенных источниках витамина B1 (Tang et al., 2010; Sañudo-Wilhelmy et al., 2014).
Недавние исследования, проведенные с использованием подходов секвенирования нового поколения и клеточных культур, продемонстрировали, что ауксотрофия витамина B1 еще более широко распространена и сложна, чем предполагалось в первоначальных отчетах.В настоящее время выясняется, что многие организмы развили пробелы в канонических путях биосинтеза витамина B1 и, следовательно, нуждаются в одном или нескольких предшественниках тиамина или продуктах деградации, далее именуемых соединениями, родственными тиамину (TRC). Наиболее распространенному гетеротрофу в океане, SAR11, не хватает ферментов для синтеза пиримидиновой части тиамина, 4-амино-5-гидроксиметил-2-метилпиримидина (HMP), и для завершения биосинтеза тиамина он должен получать HMP из растворенного пула в окружающей морской воде. Карини и др., 2014). Подобная ауксотрофия наблюдалась у некоторых гаптофитных и страминопиловых водорослей, где было замечено, что организмы отдавали предпочтение HMP и его аналогу, связанному со спасением, 4-амино-5-аминометил-2-метилпиримидину (AmMP), а не интактному тиамину (McRose et al. ., 2014; Гутовска и др., 2017). Ауксотрофия как по тиазольному, так и по пиримидиновому фрагменту наблюдалась у морских пикоэукариот. Геномные исследования показывают, что эти организмы обязательно нуждаются в экзогенных источниках как HMP, так и 4-метил-5-тиазолэтанола (HET), в то время как они поддерживают биосинтетический механизм для лигирования этих двух очищенных фрагментов для удовлетворения их метаболической потребности в тиамине (McRose et al.
Наиболее распространенному гетеротрофу в океане, SAR11, не хватает ферментов для синтеза пиримидиновой части тиамина, 4-амино-5-гидроксиметил-2-метилпиримидина (HMP), и для завершения биосинтеза тиамина он должен получать HMP из растворенного пула в окружающей морской воде. Карини и др., 2014). Подобная ауксотрофия наблюдалась у некоторых гаптофитных и страминопиловых водорослей, где было замечено, что организмы отдавали предпочтение HMP и его аналогу, связанному со спасением, 4-амино-5-аминометил-2-метилпиримидину (AmMP), а не интактному тиамину (McRose et al. ., 2014; Гутовска и др., 2017). Ауксотрофия как по тиазольному, так и по пиримидиновому фрагменту наблюдалась у морских пикоэукариот. Геномные исследования показывают, что эти организмы обязательно нуждаются в экзогенных источниках как HMP, так и 4-метил-5-тиазолэтанола (HET), в то время как они поддерживают биосинтетический механизм для лигирования этих двух очищенных фрагментов для удовлетворения их метаболической потребности в тиамине (McRose et al. , 2014; Паерл и др., 2015; Гутовска и др., 2017). Интересно, что эксперименты с клеточными культурами показывают, что эти пикоэукариоты нуждаются в HMP, но не могут использовать HET. Вместо этого они могут использовать только недавно идентифицированный отдельный предшественник тиамина, 5-(2-гидроксиэтил)-4-метил-1,3-тиазол-2-карбоновую кислоту (cHET) (Paerl et al., 2016, 2018a). Метагеномные исследования морской среды использовали закономерности в разрывах путей для выявления нескольких распространенных вариантов ауксотрофии витамина B1, что привело к выводу, что большая часть бактериопланктона в природе зависит от одного или нескольких предшественников витамина B1 или продуктов деградации (Gómez-Consarnau et al., 2018; Паэрл и др., 2018b).
, 2014; Паерл и др., 2015; Гутовска и др., 2017). Интересно, что эксперименты с клеточными культурами показывают, что эти пикоэукариоты нуждаются в HMP, но не могут использовать HET. Вместо этого они могут использовать только недавно идентифицированный отдельный предшественник тиамина, 5-(2-гидроксиэтил)-4-метил-1,3-тиазол-2-карбоновую кислоту (cHET) (Paerl et al., 2016, 2018a). Метагеномные исследования морской среды использовали закономерности в разрывах путей для выявления нескольких распространенных вариантов ауксотрофии витамина B1, что привело к выводу, что большая часть бактериопланктона в природе зависит от одного или нескольких предшественников витамина B1 или продуктов деградации (Gómez-Consarnau et al., 2018; Паэрл и др., 2018b).
Недавние результаты, рассмотренные выше, указывают на то, что пути биосинтеза витамина B1 являются местом существенной эволюционной изменчивости, связанной с взаимодействиями между клетками планктона с участием промежуточных звеньев TRC. Объяснения этой изменчивости варьируются от экономии клеток на затратах на создание соединений, доступных из окружающей среды, до более сложных интерпретаций, включающих перенос соединений в качестве основы для сложных коэволюционных процессов (например,конкуренция и мутуализм).
Первоначальным источником всех TRC в океанической растворенной фазе является микробная метаболическая активность, за которой в некоторых случаях следует внеклеточный абиотический распад (Jurgenson et al., 2009). Хотя имеются существенные доказательства многих типов ауксотрофии витамина B1, которые, вероятно, приводят к удалению TRC из растворенного пула, мало что известно о скорости или конкретных механизмах, связанных с поглощением и абиотическим разложением (Gold et al., 1966; Carlucci et al. ., 1969). Точно так же скорости и механизмы синтеза и высвобождения TRC из клеточной фазы в растворенную фазу неясны. Геномные данные указывают на то, что некоторые организмы, в том числе представители Cyanobacteria, способны к синтезу тиамина de novo (Sañudo-Wilhelmy et al., 2014). Однако неясно, каковы основные источники ТРК в морской системе. К этой сложности добавляется тот факт, что растворенные концентрации всех TRC (тиамин, HMP, AmMP, HET и cHET) не измерялись одновременно в океане. О распределении тиамина и, в меньшей степени, HMP сообщалось нечасто (Sañudo-Wilhelmy et al., 2012; Heal et al., 2014; Suffridge et al., 2017, 2018). Стало ясно, что измерения только этих соединений недостаточно, чтобы полностью понять круговорот тиамина и его связь с микробным сообществом.
О распределении тиамина и, в меньшей степени, HMP сообщалось нечасто (Sañudo-Wilhelmy et al., 2012; Heal et al., 2014; Suffridge et al., 2017, 2018). Стало ясно, что измерения только этих соединений недостаточно, чтобы полностью понять круговорот тиамина и его связь с микробным сообществом.
Здесь мы сообщаем о самых полных измерениях растворенных концентраций TRC, которые когда-либо проводились. Пять TRC, которые, как предполагается, важны для связанных с B1 взаимодействий в планктоне, были одновременно измерены на вертикальных профилях вдоль широтного разреза в северной Атлантике.Пробы были собраны во время фазы накопления весеннего цветения в Северной Атлантике в марте 2018 г. в рамках Проекта аэрозолей Северной Атлантики и экосистем штата Мэн (NAAMES) (Behrenfeld et al., 2019). Полный набор физических, химических и биологических океанографических данных, включая данные о разнообразии молекулярных сообществ, сопровождал измерения концентрации TRC. Мы решили измерить пространственные закономерности растворенных концентраций TRC, чтобы улучшить понимание роли TRC в структурировании сообществ планктона. Поскольку витамин B1 требуется многим ферментам центрального метаболизма, мы предположили, что повышенное обмеление светлых и смешанных слоев во время фазы накопления цветения приведет к усилению микробной потребности в витамине B1 и, таким образом, к истощению растворенных TRC. Кроме того, мы предположили, что темпы роста и метаболическая активность будут ниже в условиях, предшествующих цветению, что приведет к почти устойчивым концентрациям растворенных TRC, отражающим более сбалансированные скорости роста и потери в микробных сообществах.
Поскольку витамин B1 требуется многим ферментам центрального метаболизма, мы предположили, что повышенное обмеление светлых и смешанных слоев во время фазы накопления цветения приведет к усилению микробной потребности в витамине B1 и, таким образом, к истощению растворенных TRC. Кроме того, мы предположили, что темпы роста и метаболическая активность будут ниже в условиях, предшествующих цветению, что приведет к почти устойчивым концентрациям растворенных TRC, отражающим более сбалансированные скорости роста и потери в микробных сообществах.
Методы
Коллекция образцов
пробы были отобраны в рамках исследования аэрозолей и морских экосистем Северной Атлантики (NAAMES) на борту НИС Atlantis в марте и апреле 2018 года (Behrenfeld et al., 2019). Пробы были отобраны на пяти станциях вдоль примерно меридионального разреза в северо-западной Атлантике от 39 до 44° с. ш. (рис. 1). Весь отбор проб производился в светлое время суток между восходом солнца и солнечным полуднем. Для отбора проб использовали пробоотборник воды CTD-rosette с 24 10-литровыми бутылями Нискина (Seabird 911+; стандартные датчики электропроводности, температуры и давления).
Для отбора проб использовали пробоотборник воды CTD-rosette с 24 10-литровыми бутылями Нискина (Seabird 911+; стандартные датчики электропроводности, температуры и давления).
Рисунок 1 . Географический, химический и биологический контекст экспедиции NAAMES4. (A) Местоположение пяти станций показано в пределах океанографических регионов Северной Атлантики на основе средней динамической топографии, определенной Della Penna and Gaube (2019). (B–F) Глубинные профили основных биологических и химических параметров с каждой станции. Показаны только глубины, на которых также были собраны TRC. Цвет профиля соответствует цвету станции (А) .Полное представление, объяснение и анализ этих данных можно найти в Baetge et al. (2020).
Экологические данные проекта NAAMES находятся в открытом доступе SeaBASS Ocean Biology DAAC (http://seabass.gsfc.nasa.gov, doi: 10.5067/SeaBASS/NAAMES/DATA001). Образцы хлора и фосфата собирали и обрабатывали, как описано Mojica et al. (2020). Обилие бактерий, бактериальную продуктивность (включение лейцина) и растворенный органический углерод (DOC) собирали и обрабатывали, как описано Baetge et al.(2020). Измерения физической океанографии, включая среднюю динамическую топографию, были проанализированы Делла Пенна и Гаубе (2019).
(2020). Обилие бактерий, бактериальную продуктивность (включение лейцина) и растворенный органический углерод (DOC) собирали и обрабатывали, как описано Baetge et al.(2020). Измерения физической океанографии, включая среднюю динамическую топографию, были проанализированы Делла Пенна и Гаубе (2019).
Соединения, родственные тиамину
пробы растворенного TRC были собраны, как описано ранее (Suffridge et al., 2017). Вкратце, клетки и частицы удаляли из морской воды, собранной из розетки для отбора проб, путем осторожной перистальтической фильтрации (30 мл/мин) через фильтр Sterivex 0,22 мкм (мембрана PES, Millipore, Burlington, MA, USA). Один литр фильтрата собирали в промытые кислотой и промытые метанолом бутылки из полиэтилена высокой плотности янтарного цвета (Nalgene).Образцы были немедленно заморожены при температуре -20°C и отправлены в замороженном виде в Университет штата Орегон для анализа. Образцы были защищены от света на протяжении всего процесса. Из-за ограничений водного баланса отбирали только одну пробу с глубины.
TRC были извлечены из морской воды, как описано ранее (Suffridge et al., 2017). Образцы оттаивали и подкисляли до pH 6,5, используя 1M HCl. TRC экстрагировали из морской водной матрицы с использованием твердофазной экстракции со смолой Bondesil C 18 (Agilent).Образцы пропускали через 8 мл смолы со скоростью 1 мл мин -1 . Было определено, что все TRC сохранились на смоле. Затем смолу промывали водой класса LCMS для удаления солей, и TRC элюировали из колонки с использованием 12 мл метанола класса LCMS. Смесь метанол-TRC упаривали до 250 мкл, используя азотную сушилку с продувкой (Glass Col), обеспечивая коэффициент концентрации шести порядков между концентрацией образца в окружающей среде и концентрацией, анализируемой на ЖХ-МС.Затем образцы хранили при -80°C до проведения анализа ЖХ-МС.
Метод жидкостной хроматографии и масс-спектрометрии (ЖХ-МС) был разработан для одновременного измерения концентраций TRC. Анализ проводили с использованием тройного квадрупольного масс-спектрометра Applied Biosystems 4000 Q-Trap с интерфейсом ESI, соединенного с жидкостным хроматографом Shimatzu LC-20AD. Программное обеспечение Applied Biosystems Analyst и ABSciex Multiquant использовалось для работы с прибором и количественного определения образцов.Для хроматографического разделения использовали колонку Poroshell 120 PFP, 3 × 150 мм, 2,7 мкм для ВЭЖХ (Agilent) с защитной колонкой Poroshell 120 PFP, 2 × 5 мм, 2,7 мкм (Agilent). Температура колонки была изократической при 40°С. Подвижными фазами для ВЭЖХ были вода класса MS (Fisher) с 0,1% муравьиной кислоты и ацетонитрил класса MS (Fisher) с 0,1% муравьиной кислоты. Использовали 15-минутный бинарный градиент со скоростью потока 200 мкл/мин -1 и начальной концентрацией 3% ацетонитрила, повышающейся до 100% ацетонитрила за 7 минут, и повторным уравновешиванием колонки при 3% ацетонитриле в течение 6 минут.Третий насос для ВЭЖХ со скоростью потока 100 мкл/мин -1 ацетонитрила (0,1% муравьиной кислоты) был подключен к смесительной колонке с тройником для повышения эффективности ионизации, поскольку большинство ТРК элюируют из колонки в водной фазе градиента.
Программное обеспечение Applied Biosystems Analyst и ABSciex Multiquant использовалось для работы с прибором и количественного определения образцов.Для хроматографического разделения использовали колонку Poroshell 120 PFP, 3 × 150 мм, 2,7 мкм для ВЭЖХ (Agilent) с защитной колонкой Poroshell 120 PFP, 2 × 5 мм, 2,7 мкм (Agilent). Температура колонки была изократической при 40°С. Подвижными фазами для ВЭЖХ были вода класса MS (Fisher) с 0,1% муравьиной кислоты и ацетонитрил класса MS (Fisher) с 0,1% муравьиной кислоты. Использовали 15-минутный бинарный градиент со скоростью потока 200 мкл/мин -1 и начальной концентрацией 3% ацетонитрила, повышающейся до 100% ацетонитрила за 7 минут, и повторным уравновешиванием колонки при 3% ацетонитриле в течение 6 минут.Третий насос для ВЭЖХ со скоростью потока 100 мкл/мин -1 ацетонитрила (0,1% муравьиной кислоты) был подключен к смесительной колонке с тройником для повышения эффективности ионизации, поскольку большинство ТРК элюируют из колонки в водной фазе градиента.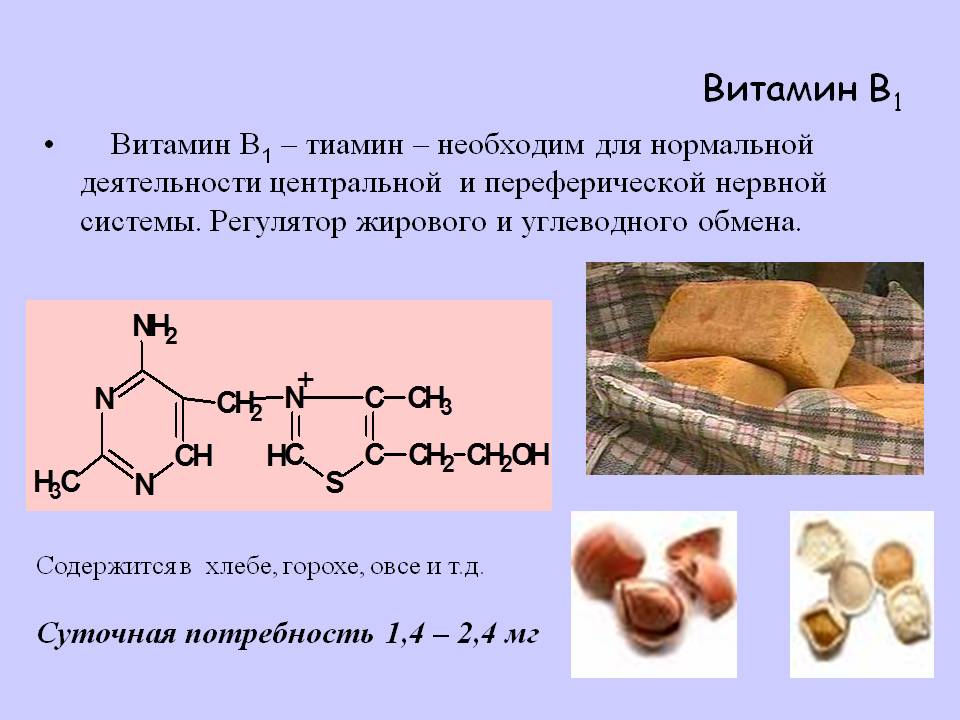 . В источнике ESI использовалось напряжение распыления 5200 В и температура источника 450°C. Давление завесного газа было установлено на уровне 30 фунтов на квадратный дюйм. Масс-спектрометр работал в режиме положительных ионов. Информация о конкретном соединении, включая параметры MRM, время удерживания на колонке и пределы обнаружения, представлена в таблице 1.Объем вводимой пробы составлял 20 мкл, пробы анализировали в трех повторностях. Образцы были без разбора рандомизированы перед анализом. Для компенсации матричных эффектов в качестве внутреннего стандарта использовали 13 C-меченый тиамин. Анализ ЖХ-МС был проведен в Центре масс-спектрометрии Университета штата Орегон.
. В источнике ESI использовалось напряжение распыления 5200 В и температура источника 450°C. Давление завесного газа было установлено на уровне 30 фунтов на квадратный дюйм. Масс-спектрометр работал в режиме положительных ионов. Информация о конкретном соединении, включая параметры MRM, время удерживания на колонке и пределы обнаружения, представлена в таблице 1.Объем вводимой пробы составлял 20 мкл, пробы анализировали в трех повторностях. Образцы были без разбора рандомизированы перед анализом. Для компенсации матричных эффектов в качестве внутреннего стандарта использовали 13 C-меченый тиамин. Анализ ЖХ-МС был проведен в Центре масс-спектрометрии Университета штата Орегон.
Таблица 1 . Специфические для соединения параметры LCMS.
Анализ данных проводился в программной среде R (R Core Team, 2015).Пакет corrplot использовался для анализа корреляций Спирмена между TRC и основными параметрами (дополнительный рисунок 1). Визуализация данных была проведена с использованием пакета ggplot (Wickham, 2009), а полученные рисунки были отредактированы в Adobe Illustrator (Adobe) для эстетики.
Экстракция ДНК и секвенирование ампликонов
Микробную биомассу собирали на фильтре Sterivex с размером пор 0,22 мкм (мембрана PES, Millipore, Burlington, MA, USA) с использованием восьмиканального перистальтического насоса (скорость потока 30 мл/мин).Фильтрат использовали для анализа TRC, как описано выше. К фильтрам добавляли один миллилитр буфера для лизиса сахарозы и хранили при -80°С. Нуклеиновые кислоты экстрагировали в соответствии с протоколом фенол:хлороформ, описанным Giovannoni et al. (1996) и Morris et al. (2005). Концентрацию ДНК измеряли с помощью анализов Quant-iT (Invitrogen, Карлсбад, Калифорния) на флюоромотерах Qubit (Invitrogen).
Амплификацию области V1-V2 гена 16S рРНК проводили с использованием праймеров 27F (5′-AGAGTTTGATCNTGGCTCAG-3) и 338 RPL (5′-GCWGCCWCCCGTAGGWGT-3′), прикрепленных к соответствующим выступающим адаптерам Illumina (Vergin et al. ., 2013; Иллюмина Инк.). Условия полимеразной цепной реакции (ПЦР) и создание библиотеки были выполнены в соответствии с протоколом ампликона NAAMES, как описано Bolanos et al. (2020б). Секвенирование с использованием платформы Illumina MiSeq (набор реагентов v.2; 2X250 Paired-End; Illumina Inc.) проводили в Центре геномных исследований и биокомпьютинга Университета штата Орегон, как описано ранее (Bolaños et al., 2020a,b).
(2020б). Секвенирование с использованием платформы Illumina MiSeq (набор реагентов v.2; 2X250 Paired-End; Illumina Inc.) проводили в Центре геномных исследований и биокомпьютинга Университета штата Орегон, как описано ранее (Bolaños et al., 2020a,b).
Анализы ампликонов генов 16S рРНК
Праймеры вырезали из демультиплексированных файлов fastq с помощью программного обеспечения CutAdapt (Martin et al., 2011) удаление фиксированного количества оснований (параметр -u). Обрезанные файлы fastq были отфильтрованы по качеству, дереплицированы и объединены с пакетом Dada2 R версии 1.2 (Callahan et al., 2016). Таблица вариантов последовательности ампликона (ASV) была построена, как в Bolaños et al. (2020б). Таксономическая принадлежность ASV была определена с использованием двухэтапного подхода (Bolaños et al., 2020b). Во-первых, с помощью команды assignTaxonomy, использующей набор данных silva_nr_v123 в пакете dada2. Во-вторых, пластидные, цианобактерии, ASV SAR11 и SAR202 были извлечены и филогенетически помещены в тщательно подобранные референсные деревья (Sudek et al.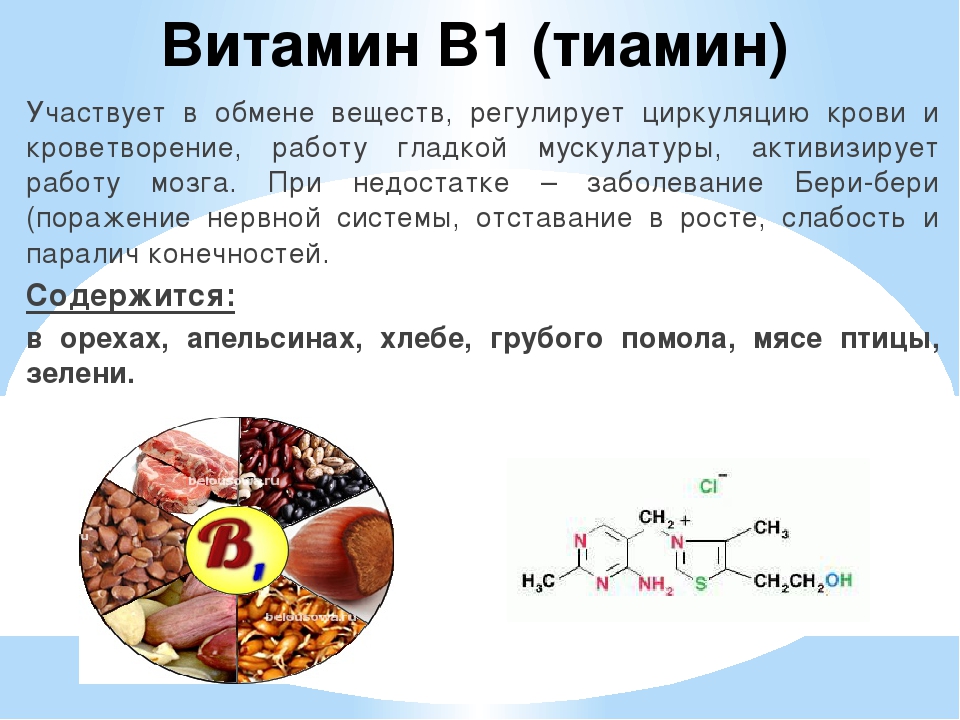 , 2015; Чой и др., 2017; Landry et al., 2017) с использованием Phyloassigner версии 089 (Vergin et al., 2013). Был построен объект phyloseq, содержащий образцы профилей станций. Отрицательные контроли брали из буферов Sucrose Lysis (SLB) и TE. Только один контрольный образец SLB (N4SLB_neg) дал результаты ампликона. Мы проанализировали распространенность потенциальных контаминационных вирусов с помощью обеззараживающей упаковки (Davis et al., 2018) и определили, что 18 вирусов из числа вирусов были получены из отрицательного контроля, поэтому они были удалены из анализа.Различия Брея-Кертиса использовались для создания ограниченной ординации (CAP) с использованием пакетов Vegan (Oksanen et al., 2007) и Phyloseq (Mcmurdie and Holmes, 2013).
, 2015; Чой и др., 2017; Landry et al., 2017) с использованием Phyloassigner версии 089 (Vergin et al., 2013). Был построен объект phyloseq, содержащий образцы профилей станций. Отрицательные контроли брали из буферов Sucrose Lysis (SLB) и TE. Только один контрольный образец SLB (N4SLB_neg) дал результаты ампликона. Мы проанализировали распространенность потенциальных контаминационных вирусов с помощью обеззараживающей упаковки (Davis et al., 2018) и определили, что 18 вирусов из числа вирусов были получены из отрицательного контроля, поэтому они были удалены из анализа.Различия Брея-Кертиса использовались для создания ограниченной ординации (CAP) с использованием пакетов Vegan (Oksanen et al., 2007) и Phyloseq (Mcmurdie and Holmes, 2013).
Дифференциальный анализ численности был выполнен с помощью DESEq2 (Love et al., 2014). Вкратце, мы классифицировали образцы на каждой станции как низкие или высокие для каждого из конгенеров B1 (рис. 5). Выбросы были исключены с использованием непараметрического числового теста на выбросы. Коэффициент, используемый для определения пределов распределения, в 3 раза превышал диапазон внутренних квартилей [нижний предел = Q1 – (3*IQR), верхний предел = Q3 + (3*IQR)].Средняя концентрация (без выбросов) каждого соединения во всех образцах использовалась в качестве порога для определения того, были ли они низкой концентрацией (ниже среднего) или высокой концентрацией (выше среднего). Каждый TRC рассматривался как независимая переменная. Для оценки распределения использовались значения TRC со всех станций и глубин. Для дифференциального анализа численности тестировались только пробы высотой более 75 м (охватывающие эвфотическую зону). Были отобраны только ASV с отсечкой альфа (padj) <0,01.Скрипты, используемые для этих анализов, можно найти по адресу https://github.com/lbolanos32/NAAMES_2020. Сценарии и таблицы, используемые для создания рисунков 4, 5, можно найти в дополнительной таблице 1 и по адресу https://github.com/lbolanos32/NAAMES_2020/tree/master/VitaminB1.
Коэффициент, используемый для определения пределов распределения, в 3 раза превышал диапазон внутренних квартилей [нижний предел = Q1 – (3*IQR), верхний предел = Q3 + (3*IQR)].Средняя концентрация (без выбросов) каждого соединения во всех образцах использовалась в качестве порога для определения того, были ли они низкой концентрацией (ниже среднего) или высокой концентрацией (выше среднего). Каждый TRC рассматривался как независимая переменная. Для оценки распределения использовались значения TRC со всех станций и глубин. Для дифференциального анализа численности тестировались только пробы высотой более 75 м (охватывающие эвфотическую зону). Были отобраны только ASV с отсечкой альфа (padj) <0,01.Скрипты, используемые для этих анализов, можно найти по адресу https://github.com/lbolanos32/NAAMES_2020. Сценарии и таблицы, используемые для создания рисунков 4, 5, можно найти в дополнительной таблице 1 и по адресу https://github.com/lbolanos32/NAAMES_2020/tree/master/VitaminB1.
Анализ ПСИ
Кодирующие белок последовательности из 31 одиночного амплифицированного генома (SAG) класса Flavobacteria были получены из базы данных IMG (https://img. jgi.doe.gov; дополнительные таблицы 2, 3) и использовались при поиске последовательностей белков. .
jgi.doe.gov; дополнительные таблицы 2, 3) и использовались при поиске последовательностей белков. .
Общедоступные SAG флавобактерий (дополнительная таблица 2), полученные из базы данных IMG, были исследованы на предмет биосинтеза и белков-транспортеров, связанных с TRC, с использованием репрезентативных профилей-HMM из базы данных PFAM (v. 28). Кроме того, профили HMM, созданные Paerl et al. (2018a) и размещены в базе данных TEED (http://www.teed.uni-stuttgart.de). Анализ последовательности по адаптированной базе данных TRC был выполнен с использованием HMMER и выходных данных для каждого домена.Совпадения, оцениваемые выше доверенных отсечений для профилей, сохранялись, и каждой белковой последовательности присваивались неперекрывающиеся HMM с наивысшей оценкой. Домены, предсказанные HMMER, сравнивали с аннотациями генов, полученными из IMG, и сохраняли конгруэнтные назначения (дополнительная таблица 4). CheckM использовали для оценки уровня полноты генома и контаминации SAG флавобактерий. Были применены два набора универсальных маркерных генов, либо специфичных для бактерий, либо для Bacteroidetes.
Были применены два набора универсальных маркерных генов, либо специфичных для бактерий, либо для Bacteroidetes.
В дополнение к ASV, полученным во время круиза NAAMES4 (Bolaños et al., 2020a), последовательности гена 16S рРНК полной длины были извлечены из 31 SAG Flavobacteria (дополнительная таблица 2). Кроме того, последовательности гена 16S рРНК были извлечены из базы данных SILVA (v.138), набора данных Северного моря (Alonso et al., 2007) и набора данных прибрежной части Северной Каролины (D’ambrosio et al., 2014) для множественных последовательностей. выравнивания с использованием MUSCLE (Эдгар, 2004). Последовательности 16S рРНК почти полной длины были обрезаны до ~ 323 п.н. с использованием Geneious Prime v. 2020.0.5 (https://www.geneious.com). Филогенетические деревья были выведены с помощью программного инструмента MAFFT с использованием алгоритма Neighbor-Joining.Модель Юкса-Кантора была применена для расчета эволюционных расстояний, а значения начальной загрузки были рассчитаны на основе 1000 повторений.
Результаты
Океанографическое распределение растворенных соединений, родственных тиамину (TRC)
Концентрации пяти родственных тиамину соединений (TRC) [то есть тиамина (B1), HMP, AmMP, HET и cHET] были измерены в растворенной фазе на пяти станциях во время экспедиции NASA NAAMES4 в марте 2018 г. (рис. 1). Все ТРК были обнаружены на каждой станции северо-западной Атлантики (табл. 2).На всех станциях и глубинах растворенные концентрации варьировались между фемптомолярными и пикомолярными уровнями [7,35–353 пМ B1, 0,09–3,45 пМ HMP, 1,76–113,5 пМ AmMP, 0,03–30,8 пМ HET и 3,76–145 пМ cHET] (таблица 2, рисунок). 2). Мы наблюдали существенную изменчивость между станциями в вертикальном распределении TRC (рис. 3) и диапазонах концентраций (рис. 2). Вероятно, это связано с различными физическими, химическими и биологическими характеристиками каждой станции (рис. 1) (Behrenfeld et al., 2019; Della Penna and Gaube, 2019).Об этом свидетельствует существенный разброс глубин смешанных слоев на пяти станциях: от 5,9 м на станции 2. 1 до 214 м на станции 3 (Fox et al., 2020).
1 до 214 м на станции 3 (Fox et al., 2020).
Таблица 2 . Концентрации растворенных ТРК в пикомолях на всех станциях и глубинах.
Рисунок 2 . Гистограммы концентраций растворенных родственных тиамину соединений (TRC) на каждой станции показывают существенные различия в диапазонах концентраций между станциями. Пикомолярные концентрации каждого TRC нанесены на график в масштабе log10.Наибольшие концентрации ТРК наблюдались на станции 2.
Рисунок 3 . Распределение по глубине растворенного соединения, родственного тиамину (TRC). Вертикальные распределения нанесены на график с использованием отдельной оси, чтобы подчеркнуть сходство в тенденциях распределения TRC. Тиамин (B1) показан черным цветом, TRC пиримидина (HMP и AmMP) — зеленым, а TRC тиазола (HET и cHET) — фиолетовым. Цвет оси относится к цвету профиля глубины.Баллы и планки погрешностей представляют собой средние значения и стандартные отклонения технических повторов ( n = 3).
Вертикальное распределение TRC часто определялось обеднением приповерхностных и глубинных проб (>150 м) по сравнению с обогащением на средних глубинах (~100 м; рис. 3). Например, все TRC на станции 1 имеют одинаковое вертикальное распределение, при этом низкие уровни у поверхности увеличиваются до максимальной концентрации на глубине 75 м (рис. 3). Медиана концентрации TRC на станции 1 составила 26.65 пМ для B1, 0,17 пМ для HMP, 6,44 пМ для AmMP, 0,08 пМ для HET и 17,93 пМ для cHET (рис. 2). Бактериальная продукция, численность бактерий и максимумы хлорофилла а (Chl а) также были на высоте 75 м, и профили этих свойств положительно коррелировали с профилями TRC (рис. 1, дополнительная рис. 1). Глубина перемешанного слоя на этой станции составляла 80 м, что совпадает с максимальным значением TRC и бактериальной продукции, предполагая, что на этой станции повышенная скорость метаболизма приводит к тому, что скорость накопления TRC в растворенном пуле превышает скорость смешивания вблизи дно смешанного слоя.
Наибольшие наблюдаемые концентрации TRC были на станции 2 (глубина смешанного слоя составляла 65 м), со средними значениями 51,05 пМ для B1, 0,71 пМ для HMP, 21,15 пМ для AmMP, 2,12 пМ для HET и 77,78 пМ для cHET ( Фигура 2). На этой станции все профили TRC демонстрировали одинаковые вертикальные профили, которые оставались постоянными в пределах перемешанного слоя, а затем увеличивались от нижней части перемешанного слоя до 150 м (рис. 3). В отличие от станции 1, на станции 2 наблюдались сильные отрицательные корреляции между концентрациями TRC и бактериальной продукцией, численностью бактерий, DOC и Chl a, все из которых были на самых высоких значениях для круиза (рис. 1, дополнительная рис. 1).Кроме того, все TRC на станции 2 положительно коррелировали с фосфатом.
Аналогичные тенденции в распределении TRC наблюдались на станции 2.1, хотя амплитуда была значительно ниже, со средними концентрациями TRC 13,15 пМ для B1, 0,23 пМ для HMP, 6,30 пМ для AmMP, 0,27 для HET и 6,78 пМ для cHET (рис. 2). Разрешение по глубине в верхних 300 м на станции 2.1 было меньше, чем на станции 2, при этом были отобраны только три глубины (рис. 3). Концентрации ТРК в двух глубоких пробах, отобранных на глубине 1000 и 2600 м, были такими же или ниже самых низких концентраций ТРК в верхних 300 м на станции 2.1 (табл. 2). Все TRC были положительно коррелированы с минимумами на 25 и 300 м по сравнению с их максимальной концентрацией на 100 м (рис. 3, дополнительная рис. 1). HET разделяет минимум 2 м с другими TRC, но имеет максимум 300 м вместо 100 м. Глубина перемешанного слоя на этой станции составляла 6 м, и все пробы ТРК были отобраны ниже этой глубины. В верхних 300 м концентрации TRC сильно положительно коррелировали с концентрациями фосфатов. Все TRC слабо отрицательно коррелировали с бактериальной продукцией, численностью бактерий, Chl a и DOC.
2). Разрешение по глубине в верхних 300 м на станции 2.1 было меньше, чем на станции 2, при этом были отобраны только три глубины (рис. 3). Концентрации ТРК в двух глубоких пробах, отобранных на глубине 1000 и 2600 м, были такими же или ниже самых низких концентраций ТРК в верхних 300 м на станции 2.1 (табл. 2). Все TRC были положительно коррелированы с минимумами на 25 и 300 м по сравнению с их максимальной концентрацией на 100 м (рис. 3, дополнительная рис. 1). HET разделяет минимум 2 м с другими TRC, но имеет максимум 300 м вместо 100 м. Глубина перемешанного слоя на этой станции составляла 6 м, и все пробы ТРК были отобраны ниже этой глубины. В верхних 300 м концентрации TRC сильно положительно коррелировали с концентрациями фосфатов. Все TRC слабо отрицательно коррелировали с бактериальной продукцией, численностью бактерий, Chl a и DOC.
В отличие от всех других станций, на станции 3 наблюдалось поверхностное обогащение всех TRC (кроме HET) (рис. 3). Ниже наблюдаемых поверхностных максимумов все концентрации TRC оставались низкими и относительно постоянными. Средние концентрации TRC на станции 3 составляли 11,45 пМ для B1, 0,21 пМ для HMP, 5,89 пМ для AmMP, 0,66 пМ для HET и 9,45 пМ для cHET (рис. 2). HET положительно коррелировал с фосфатом и отрицательно коррелировал с другими TRC, а также биологическими и химическими параметрами (дополнительная фигура 1).HMP положительно коррелировал с фосфатом, слабо отрицательно коррелировал с обилием бактерий и Chl a. Другие TRC отрицательно коррелировали с фосфатом и положительно коррелировали с численностью бактерий, бактериальной продукцией, Chl a и DOC. Глубина перемешанного слоя на станции 3 была определена равной 214 м, что является максимальным значением, наблюдавшимся в нашей выборке.
Средние концентрации TRC на станции 3 составляли 11,45 пМ для B1, 0,21 пМ для HMP, 5,89 пМ для AmMP, 0,66 пМ для HET и 9,45 пМ для cHET (рис. 2). HET положительно коррелировал с фосфатом и отрицательно коррелировал с другими TRC, а также биологическими и химическими параметрами (дополнительная фигура 1).HMP положительно коррелировал с фосфатом, слабо отрицательно коррелировал с обилием бактерий и Chl a. Другие TRC отрицательно коррелировали с фосфатом и положительно коррелировали с численностью бактерий, бактериальной продукцией, Chl a и DOC. Глубина перемешанного слоя на станции 3 была определена равной 214 м, что является максимальным значением, наблюдавшимся в нашей выборке.
Станция 4 была единственной станцией, на которой вертикальное распределение TRC не коррелировалось. На рисунке 3 можно увидеть три различных модели распределения TRC.Распределения HET и HMP имеют сильную положительную корреляцию, демонстрируя истощение на 0–25 м по сравнению с максимумами на 75 м, минимумами на 150 м и максимумами на 300 м (рис. 3, дополнительная рис. 1). B1 и AmMP положительно коррелируют и истощаются на глубине 0–75 м по сравнению с глубиной 150 м и ниже (рис. 3). cHet обогащен на поверхности относительно 75 м (рис. 3). Средние концентрации TRC составляли 13,4 пМ для B1, 0,18 пМ для HMP, 8,50 пМ для AmMP, 0,15 пМ для HET и 7,41 пМ для cHET (рис. 2). TRC, за исключением cHET, отрицательно коррелировали с бактериальной продукцией, численностью бактерий, Chl a, DOC (за исключением B1, который имеет положительную корреляцию) и фосфатом.Напротив, cHET положительно коррелировал со всеми основными параметрами, кроме фосфата, где он имел отрицательную корреляцию (рис. 1). Глубина перемешанного слоя на этой станции составляла 128 м, поэтому большинство различий в наблюдаемых закономерностях распределения имело место в перемешанном слое.
3, дополнительная рис. 1). B1 и AmMP положительно коррелируют и истощаются на глубине 0–75 м по сравнению с глубиной 150 м и ниже (рис. 3). cHet обогащен на поверхности относительно 75 м (рис. 3). Средние концентрации TRC составляли 13,4 пМ для B1, 0,18 пМ для HMP, 8,50 пМ для AmMP, 0,15 пМ для HET и 7,41 пМ для cHET (рис. 2). TRC, за исключением cHET, отрицательно коррелировали с бактериальной продукцией, численностью бактерий, Chl a, DOC (за исключением B1, который имеет положительную корреляцию) и фосфатом.Напротив, cHET положительно коррелировал со всеми основными параметрами, кроме фосфата, где он имел отрицательную корреляцию (рис. 1). Глубина перемешанного слоя на этой станции составляла 128 м, поэтому большинство различий в наблюдаемых закономерностях распределения имело место в перемешанном слое.
Взаимосвязь между TRC и составом микробного сообщества
Взаимосвязь между TRC, параметрами окружающей среды и составом микробного сообщества исследовали с помощью канонического анализа главных координат (CAP; рис. 4).Мы создали профили ампликона с высокой пропускной способностью из твердых частиц, собранных на фильтрах, используемых для получения растворенных образцов TRC. Состав сообщества был проанализирован до уровня ASV для создания профилей бактериопланктона и фитопланктона (цианобактерии и эукариотические пластиды) с высоким разрешением (Bolaños et al., 2020a,b). Профили ASV демонстрируют два основных паттерна ограниченной ординации. Во-первых, кластеры выборок, соответствующие каждой станции, следуют градиенту несходства по первому компоненту, который объясняет большую часть дисперсии (39.8%). Эта организация предполагает различие в составе сообщества станций по долготе. Пробы со станций 1, 2.1 и 3 сгруппированы вместе (западная группа), а со станций 2 и 4 (восточная группа) обособлены и образуют отдельные кластеры (рис. 1). Дифференциацию станций-кластеров по второй оси можно объяснить их широтой, так как северные станции имеют более положительное положение по оси, чем южные. Во-вторых, состав сообщества в пробах выше 75 м (охватывающих эвфотическую зону) был плотно сгруппирован на всех станциях, кроме станции 2, где наблюдались самые высокие наблюдаемые концентрации ТРК и биологические параметры (рис.
4).Мы создали профили ампликона с высокой пропускной способностью из твердых частиц, собранных на фильтрах, используемых для получения растворенных образцов TRC. Состав сообщества был проанализирован до уровня ASV для создания профилей бактериопланктона и фитопланктона (цианобактерии и эукариотические пластиды) с высоким разрешением (Bolaños et al., 2020a,b). Профили ASV демонстрируют два основных паттерна ограниченной ординации. Во-первых, кластеры выборок, соответствующие каждой станции, следуют градиенту несходства по первому компоненту, который объясняет большую часть дисперсии (39.8%). Эта организация предполагает различие в составе сообщества станций по долготе. Пробы со станций 1, 2.1 и 3 сгруппированы вместе (западная группа), а со станций 2 и 4 (восточная группа) обособлены и образуют отдельные кластеры (рис. 1). Дифференциацию станций-кластеров по второй оси можно объяснить их широтой, так как северные станции имеют более положительное положение по оси, чем южные. Во-вторых, состав сообщества в пробах выше 75 м (охватывающих эвфотическую зону) был плотно сгруппирован на всех станциях, кроме станции 2, где наблюдались самые высокие наблюдаемые концентрации ТРК и биологические параметры (рис. 1, 3).Сообщества ниже 75 метров отличаются от сообществ эвфотической зоны. Сдвиг сообщества на 75 м в основном объясняется ВПП фитопланктона (эукариотические пластиды и цианобактерии). Состав сообщества на южных станциях коррелировал с более высокой бактериальной продуктивностью, численностью и флуоресценцией. Более высокие концентрации фосфатов были связаны с восточно-мезопелагическими образцами, а более высокие концентрации DOC отличались от эвфотических западных образцов. TRC были переменными, которые в наименьшей степени способствовали объяснению различий между сообществами в линейной модели.Однако концентрации TRC демонстрировали специфические ассоциации с элементами структуры сообщества, о чем свидетельствуют различия в направлениях стрелок. Это наблюдение предполагает, что отдельные подмножества организмов в микробных сообществах могут быть ответственны за растворенные концентрации TRC или реагировать на них (рис. 4).
1, 3).Сообщества ниже 75 метров отличаются от сообществ эвфотической зоны. Сдвиг сообщества на 75 м в основном объясняется ВПП фитопланктона (эукариотические пластиды и цианобактерии). Состав сообщества на южных станциях коррелировал с более высокой бактериальной продуктивностью, численностью и флуоресценцией. Более высокие концентрации фосфатов были связаны с восточно-мезопелагическими образцами, а более высокие концентрации DOC отличались от эвфотических западных образцов. TRC были переменными, которые в наименьшей степени способствовали объяснению различий между сообществами в линейной модели.Однако концентрации TRC демонстрировали специфические ассоциации с элементами структуры сообщества, о чем свидетельствуют различия в направлениях стрелок. Это наблюдение предполагает, что отдельные подмножества организмов в микробных сообществах могут быть ответственны за растворенные концентрации TRC или реагировать на них (рис. 4).
Рисунок 4 . Канонический анализ основных координат (CAP) профилей 16S рРНК ASV, ограниченный физическими и химическими параметрами окружающей среды, включая измерения TRC. Ординация была построена с помощью расстояний Брея-Кертиса между выборками. Форма точек указывает на глубину происхождения проб, а цвета обозначают станции, на которых они были собраны. На оси представлены первый и второй ограниченные компоненты, а проценты соответствуют доле дисперсии, объясненной линейной комбинацией выбранных параметров окружающей среды. Стрелки показывают направление отклонения каждого параметра окружающей среды.
Ординация была построена с помощью расстояний Брея-Кертиса между выборками. Форма точек указывает на глубину происхождения проб, а цвета обозначают станции, на которых они были собраны. На оси представлены первый и второй ограниченные компоненты, а проценты соответствуют доле дисперсии, объясненной линейной комбинацией выбранных параметров окружающей среды. Стрелки показывают направление отклонения каждого параметра окружающей среды.
Возможные связи между распределением TRC и составом микробного сообщества в толще воды
Для дальнейшего изучения общего влияния концентрации TRC на состав микробного сообщества во всей толще воды мы сначала классифицировали образцы, которые были ниже общего среднего значения для каждого TRC, как «низкие», а выше среднего — как «высокие» (рис. 5). .Для определения этих интервалов в качестве порога использовалось среднее значение (исключая выбросы) концентрации каждого TRC во всех образцах на всех станциях (22,27 пМ B1, 0,28 пМ HMP, 10,89 пМ AmMP, 0,47 пМ HET и 11,48 пМ cHET). На некоторых станциях разные конгенеры имели одинаковую ковариацию от высокого к низкому (дополнительный рисунок 1). Мы протестировали среднее геометрическое нормированных изменений численности ASV между обоими условиями на каждой станции для каждой возможной картины классифицированных комбинаций TRC. Наибольшее количество различий ASV между низкими и высокими бинами наблюдалось для B1 на станциях 1 и 2 (рис. 5, дополнительная таблица 1).Мы не обнаружили обогащения при сравнении ASV, связанного с изменениями концентрации других TRC на этих станциях. Меньшее количество ASV представило различия в их численности на станции 3 и 4. На станции 3 семь ASV были дифференциально многочисленны: три реагировали на ковариацию B1, cHET и HMP, а четыре — на HET. На станции 4 пять ASV демонстрируют дифференциальную численность по сравнению с ковариацией HET и AmMP (дополнительная таблица 1). Меньшее количество откликов биологических сигналов (в виде дифференциальной численности ASV) на станциях 3 и 4 можно объяснить тем, что концентрации TRC проявляют меньшую изменчивость на этих станциях в толще воды (рис.
На некоторых станциях разные конгенеры имели одинаковую ковариацию от высокого к низкому (дополнительный рисунок 1). Мы протестировали среднее геометрическое нормированных изменений численности ASV между обоими условиями на каждой станции для каждой возможной картины классифицированных комбинаций TRC. Наибольшее количество различий ASV между низкими и высокими бинами наблюдалось для B1 на станциях 1 и 2 (рис. 5, дополнительная таблица 1).Мы не обнаружили обогащения при сравнении ASV, связанного с изменениями концентрации других TRC на этих станциях. Меньшее количество ASV представило различия в их численности на станции 3 и 4. На станции 3 семь ASV были дифференциально многочисленны: три реагировали на ковариацию B1, cHET и HMP, а четыре — на HET. На станции 4 пять ASV демонстрируют дифференциальную численность по сравнению с ковариацией HET и AmMP (дополнительная таблица 1). Меньшее количество откликов биологических сигналов (в виде дифференциальной численности ASV) на станциях 3 и 4 можно объяснить тем, что концентрации TRC проявляют меньшую изменчивость на этих станциях в толще воды (рис. 2).
2).
Рисунок 5 . Сравнение океанографической и микробной динамики на станциях 1 и 2. (A) Глубинные профили растворенного B1 (пМ, черный), бактериальной продукции (пмоль Leu/л/ч, бирюзовый) и фосфата (мкМ, коричневый) на станциях 1 и 2. (B) Тепловая карта дифференциально распространенных ASV на глубинах с высокой концентрацией B1 (желтые ореолы A ) по сравнению с низкой концентрацией B1 (зеленые ореолы A ). Красным цветом показаны положительные логарифмические 2-кратные изменения, указывающие на высокую численность ASV на глубинах с низким B1 по сравнению с высоким B1.Синим цветом показаны отрицательные log2-кратные изменения, указывающие на высокую численность ASV на глубинах с высоким B1 по сравнению с низким B1. Наносятся на график ASV с дифференциальной численностью (логарифмическое 2-кратное изменение больше 2 и меньше -2) между образцами B1 с высокой и низкой концентрацией на станциях 1 и 2. Для этого анализа рассматривались только образцы выше 75 метров. Таксономическая принадлежность нанесенных на график ASV показана по оси ординат. Полную таксономическую информацию можно найти в дополнительной таблице 1. (C) Филогенетические взаимоотношения последовательностей генов 16S рРНК из NAAMES4 Flavobacteria ASV, обильные на расстоянии 5–25 м на станции 2 (красные квадраты, тип столбца), общедоступные амплификации одной клетки Flavobacteria геномы (SAG; серые квадраты, шрифтовая колонка) и ASV Bacteroidetes (белые квадраты, типовая колонка), извлеченные из прибрежных районов Северного моря (Alonso et al., 2007) и прибрежный бактериопланктон Северной Каролины (D’ambrosio et al., 2014). Общедоступные SAG флавобактерий (дополнительные таблицы 2–4) были запрошены для подтверждения способности синтеза B1. Геномы, в которых была обнаружена способность к синтезу B1, помечены бирюзовыми квадратами в столбце ауксотрофов B1, а черные квадраты указывают на предполагаемую ауксотрофность B1 (дополнительная таблица 4).
Для этого анализа рассматривались только образцы выше 75 метров. Таксономическая принадлежность нанесенных на график ASV показана по оси ординат. Полную таксономическую информацию можно найти в дополнительной таблице 1. (C) Филогенетические взаимоотношения последовательностей генов 16S рРНК из NAAMES4 Flavobacteria ASV, обильные на расстоянии 5–25 м на станции 2 (красные квадраты, тип столбца), общедоступные амплификации одной клетки Flavobacteria геномы (SAG; серые квадраты, шрифтовая колонка) и ASV Bacteroidetes (белые квадраты, типовая колонка), извлеченные из прибрежных районов Северного моря (Alonso et al., 2007) и прибрежный бактериопланктон Северной Каролины (D’ambrosio et al., 2014). Общедоступные SAG флавобактерий (дополнительные таблицы 2–4) были запрошены для подтверждения способности синтеза B1. Геномы, в которых была обнаружена способность к синтезу B1, помечены бирюзовыми квадратами в столбце ауксотрофов B1, а черные квадраты указывают на предполагаемую ауксотрофность B1 (дополнительная таблица 4). Качество генома SAG указано в столбце качества: высокое качество (> 90% полноты, < 5% загрязнения) зеленым цветом, среднее качество (> 65% полноты, < 10% загрязнения) фиолетовым и низкое качество (< 65% полнота, загрязнение <10%) выделены оранжевым цветом (дополнительная таблица 3).Все метки узлов, методы, использованные для анализа, и подробные результаты представлены на дополнительном рисунке 2, в дополнительных таблицах 2–4.
Качество генома SAG указано в столбце качества: высокое качество (> 90% полноты, < 5% загрязнения) зеленым цветом, среднее качество (> 65% полноты, < 10% загрязнения) фиолетовым и низкое качество (< 65% полнота, загрязнение <10%) выделены оранжевым цветом (дополнительная таблица 3).Все метки узлов, методы, использованные для анализа, и подробные результаты представлены на дополнительном рисунке 2, в дополнительных таблицах 2–4.
Связь концентраций витамина B1 с ASV на станциях 1 и 2 отличается от наблюдений на станциях 3 и 4 (рис. 5). Станции 1 и 2 имеют аналогичное вертикальное распределение B1 между 0 и 75 м, но концентрации на станции 2 примерно вдвое выше, чем на станции 1 (рис. 2, 3). Однако взаимосвязь между распределением ОКП и как биологическими (бактериальная продукция, обилие и Хл а), так и химическими (фосфат) параметрами для этих двух станций противоположна (рис. 5).На станции 1 мы обнаружили 17 ASV, демонстрирующих различную численность между низкими и высокими образцами. Пятнадцать из них, в том числе обильные диатомовые водоросли и шесть ASV SAR11, были обогащены образцами с высоким B1 (25 и 75 м) по сравнению с образцом с низким 5 м. На станции 2 мы обнаружили 25 ASV, демонстрирующих различную численность между низкими и высокими образцами. В отличие от станции 1, участок с низкой концентрацией В1 (5 и 25 м) был обогащен (по сравнению с сообществом 75 м) в основном копиотрофными АСВ, относящимися к Flavobacteriales (13 АСВ), Oceanospirillales (6 АСВ) и Rhodobacterales (3 АСВ). ).
Пятнадцать из них, в том числе обильные диатомовые водоросли и шесть ASV SAR11, были обогащены образцами с высоким B1 (25 и 75 м) по сравнению с образцом с низким 5 м. На станции 2 мы обнаружили 25 ASV, демонстрирующих различную численность между низкими и высокими образцами. В отличие от станции 1, участок с низкой концентрацией В1 (5 и 25 м) был обогащен (по сравнению с сообществом 75 м) в основном копиотрофными АСВ, относящимися к Flavobacteriales (13 АСВ), Oceanospirillales (6 АСВ) и Rhodobacterales (3 АСВ). ).
B1 Ауксотрофные и прототрофные типы дифференцированно распространенных линий ASV
Учитывая преобладание Flavobacteria и других копиотрофов в поверхностных водах, обедненных B1, на станции 2 (рис. 5), мы дополнительно исследовали их потенциальный статус ауксотропии B1, чтобы определить, есть ли у них потребность в TRC. С этой целью мы провели поиск в доступных базах данных и определили 31 SAG (дополнительная таблица 2) класса Flavobacteria и исследовали ключевые ферменты, участвующие в пути de novo для синтеза витамина B1 ( dxs, thiH, thiD, thiC, thiE , ( thiN , TPK1, THI80), thiG и thiL ; дополнительная таблица 4). В общей сложности 6 высококачественных SAG (завершение генома> 90%) были предполагаемыми прототрофами B1, кодирующими все ключевые ферменты для синтеза de novo B1 ( thiC, thiE и thiG ). В одном SAG (Flavobacterium sp. SG815) отсутствовали thiC и thiG , но имелось thiE и thiD , что может указывать на двойную ауксотрофию B1 и зависимость от экзогенных источников предшественников пиримидина и тиазола. Дополнительные доказательства ауксотрофии B1 у Flavobacteria были обнаружены при исследовании двух высококачественных SAG (SAT105, SCGC AAA160-P02), в которых отсутствуют все основные гены биосинтеза B1.В зависимости от набора маркерных генов оценки полноты генома выбранных SAG варьировались от 34–72% (бактерии) до 21–72% (Bacteroidetes) соответственно, в зависимости от набора маркерных генов (рис. 5, дополнительная таблица 3). Остальные SAG среднего и низкого качества с оценочной полнотой генома 23–35% не кодировали основные гены биосинтеза B1, однако остаются неубедительными в отношении их потребностей в B1 из-за их неполного характера (рис.
В общей сложности 6 высококачественных SAG (завершение генома> 90%) были предполагаемыми прототрофами B1, кодирующими все ключевые ферменты для синтеза de novo B1 ( thiC, thiE и thiG ). В одном SAG (Flavobacterium sp. SG815) отсутствовали thiC и thiG , но имелось thiE и thiD , что может указывать на двойную ауксотрофию B1 и зависимость от экзогенных источников предшественников пиримидина и тиазола. Дополнительные доказательства ауксотрофии B1 у Flavobacteria были обнаружены при исследовании двух высококачественных SAG (SAT105, SCGC AAA160-P02), в которых отсутствуют все основные гены биосинтеза B1.В зависимости от набора маркерных генов оценки полноты генома выбранных SAG варьировались от 34–72% (бактерии) до 21–72% (Bacteroidetes) соответственно, в зависимости от набора маркерных генов (рис. 5, дополнительная таблица 3). Остальные SAG среднего и низкого качества с оценочной полнотой генома 23–35% не кодировали основные гены биосинтеза B1, однако остаются неубедительными в отношении их потребностей в B1 из-за их неполного характера (рис. 5, дополнительная таблица 3). ).
5, дополнительная таблица 3). ).
Затем мы искали связи между геномами SAG, проанализированными на полноту пути, и известными ASV в наших профилях сообщества (рис. 5).В частности, чтобы выяснить, относятся ли флавобактерии, обнаруженные на станции 2, к ауксотрофам или прототрофам B1, мы построили эталонное дерево восстановленных последовательностей NAAMES ASV и гена 16S рРНК, полученных из SAG флавобактерий, набора данных Северного моря (Alonso et al., 2007). , и набор данных прибрежной Северной Каролины (D’ambrosio et al., 2014). ASV со станции 2 связаны с некультивируемыми морскими ветвями NS2(b), NS4 (согласно определению Alonso et al., 2007) и «некультивируемыми флавобактериями Атлантического океана» (рис. 5, дополнительная рис. 2).Эталонные SAG, которые группировались вместе с этими морскими кладами, либо кодировали thiL (AAA536-G18, AG-487_A19), либо не содержали каких-либо ферментов синтеза ядра B1 (JGI 02_I17, AG-485_N06). Примечательно, что последовательности генов 16S рРНК предполагаемых прототрофных SAG B1 сгруппированы вместе с морскими кладами NS3b и NS5 Flavobacteria (рис. 5). Важно отметить, что только один SAG, в котором отсутствовали ферменты синтеза ядра B1, оценивается как высокое качество, а остальные оцениваются как среднее и низкое качество (рис. 5).Следовательно, хотя эти данные свидетельствуют о том, что кластеры SAG с ампликонами NAAMES лишены способности к синтезу B1, эти результаты не являются окончательными. Несмотря на эти ограничения, эти результаты дают правдоподобную гипотезу об истощении B1 многочисленными флавобактериями вблизи поверхности на станции 2.
5). Важно отметить, что только один SAG, в котором отсутствовали ферменты синтеза ядра B1, оценивается как высокое качество, а остальные оцениваются как среднее и низкое качество (рис. 5).Следовательно, хотя эти данные свидетельствуют о том, что кластеры SAG с ампликонами NAAMES лишены способности к синтезу B1, эти результаты не являются окончательными. Несмотря на эти ограничения, эти результаты дают правдоподобную гипотезу об истощении B1 многочисленными флавобактериями вблизи поверхности на станции 2.
Обсуждение
Динамические и изменчивые океанографические условия на разрезе NAAMES4 предоставили уникальную возможность изучить, как наличие TRC влияет на микробные взаимодействия и, следовательно, на структуру сообщества.Весеннее цветение в Северной Атлантике является крупнейшим ежегодно повторяющимся цветением фитопланктона на Земле. Экспедиция NAAMES4 была запланирована для изучения фазы накопления гипотезы восстановления нарушений (DRH) для формирования весеннего цветения (Behrenfeld et al. , 2019). В DRH фаза накопления описывается как период между окончанием конвективного зимнего перемешивания и кульминацией цветения, когда рост фитопланктона ускоряется (скорость деления превышает скорость потери), а обмеление смешанных слоев и усиление солнечного света вызывают увеличение популяций фитопланктона (Беренфельд и Босс, 2014, 2018).В период накопления биомассы, когда проводились наблюдения, средние интегрированные по глубине скорости деления фитопланктона и чистая первичная продукция для поверхностного перемешанного слоя составляли 0,46 ± 0,26 сут -1 и 920 ± 405 мг С м -2 сут — 1 соответственно (Fox et al., 2020). В проекте NAAMES использовался широтный разрез для захвата различных периодов годового цикла фитопланктона во время одного рейса из-за задержки сезонных изменений с увеличением широты (рис. 1) (Behrenfeld et al., 2019). Таким образом, этот рейс охватил большую часть субтропической части северной части Атлантического океана, фиксируя биологическую и физическую изменчивость во время фазы накопления цветения (Behrenfeld et al. , 2019).
, 2019). В DRH фаза накопления описывается как период между окончанием конвективного зимнего перемешивания и кульминацией цветения, когда рост фитопланктона ускоряется (скорость деления превышает скорость потери), а обмеление смешанных слоев и усиление солнечного света вызывают увеличение популяций фитопланктона (Беренфельд и Босс, 2014, 2018).В период накопления биомассы, когда проводились наблюдения, средние интегрированные по глубине скорости деления фитопланктона и чистая первичная продукция для поверхностного перемешанного слоя составляли 0,46 ± 0,26 сут -1 и 920 ± 405 мг С м -2 сут — 1 соответственно (Fox et al., 2020). В проекте NAAMES использовался широтный разрез для захвата различных периодов годового цикла фитопланктона во время одного рейса из-за задержки сезонных изменений с увеличением широты (рис. 1) (Behrenfeld et al., 2019). Таким образом, этот рейс охватил большую часть субтропической части северной части Атлантического океана, фиксируя биологическую и физическую изменчивость во время фазы накопления цветения (Behrenfeld et al. , 2019).
Витамин B1 и связанные с ним TRC представляют собой, казалось бы, превосходный пример соединений, которые передаются микроорганизмам и могут быть основными причинами корреляций, которые используются для построения моделей планктонных сетей. B1 повсеместно необходим для катаболического и анаболического метаболизма углерода, однако большинство фитопланктона и бактериопланктона неспособны синтезировать этот кофермент de novo и для своего роста нуждаются в экзогенном источнике одного или нескольких TRC (Jurgenson et al., 2009; Санудо-Вильгельми и др., 2014 г.; Гомес-Консарнау и др., 2018 г.; Паэрл и др., 2018b). Различия в требованиях к TRC среди таксонов, которые во многих случаях могут быть оценены с достаточной точностью по геномным данным, потенциально могут формировать структуры микробных сообществ.
Океанографическое распределение растворенных соединений, родственных тиамину (TRC)
Об океанографическом распределении растворенных AmMP, cHET и HET никогда ранее не сообщалось, и полный набор TRC никогда ранее не измерялся напрямую и одновременно в океане. Здесь мы наблюдали, что концентрации этих соединений варьировались от фемптомолярных до пикомолярных со значениями 7,35–353 пМ для B1, 0,09–3,45 пМ для HMP, 1,76–113,5 пМ для AmMP, 0,03–30,8 пМ для HET и 3,76–145 пМ. для ХЭТ (табл. 2). Для наших наблюдений за тиамином (B1) мы обнаружили межквартильный диапазон (IQR) концентраций 12,0–26,7 пМ и среднюю концентрацию 16,5 пМ (таблица 2). Эти значения согласуются с предыдущими наблюдениями. В частности, ок. Сообщается о 300 дискретных измерениях растворенного B1 для океана, IQR равен 17.4–81,1 пМ при медиане 33,9 пМ (Carlucci, 1970; Carlucci and Bowes, 1972; Okbamichael and Sañudo-Wilhelmy, 2005; Koch et al., 2012, 2013; Sañudo-Wilhelmy et al., 2012, 2014; Carini et al., 2014; Heal et al., 2014; Monteverde et al., 2015; Suffridge et al., 2017, 2018). Диапазоны концентраций, которые мы наблюдали для HMP, IQR 0,16–0,39 пМ, ниже, чем ранее наблюдаемые концентрации HMP (IQR 1,53–7,02 пМ). Однако сообщалось только о 60 дискретных измерениях растворенных ГМП в океане, и все они были получены в олиготрофных регионах (Carini et al.
Здесь мы наблюдали, что концентрации этих соединений варьировались от фемптомолярных до пикомолярных со значениями 7,35–353 пМ для B1, 0,09–3,45 пМ для HMP, 1,76–113,5 пМ для AmMP, 0,03–30,8 пМ для HET и 3,76–145 пМ. для ХЭТ (табл. 2). Для наших наблюдений за тиамином (B1) мы обнаружили межквартильный диапазон (IQR) концентраций 12,0–26,7 пМ и среднюю концентрацию 16,5 пМ (таблица 2). Эти значения согласуются с предыдущими наблюдениями. В частности, ок. Сообщается о 300 дискретных измерениях растворенного B1 для океана, IQR равен 17.4–81,1 пМ при медиане 33,9 пМ (Carlucci, 1970; Carlucci and Bowes, 1972; Okbamichael and Sañudo-Wilhelmy, 2005; Koch et al., 2012, 2013; Sañudo-Wilhelmy et al., 2012, 2014; Carini et al., 2014; Heal et al., 2014; Monteverde et al., 2015; Suffridge et al., 2017, 2018). Диапазоны концентраций, которые мы наблюдали для HMP, IQR 0,16–0,39 пМ, ниже, чем ранее наблюдаемые концентрации HMP (IQR 1,53–7,02 пМ). Однако сообщалось только о 60 дискретных измерениях растворенных ГМП в океане, и все они были получены в олиготрофных регионах (Carini et al. , 2014; Суффридж и др., 2017, 2018). Наша максимальная наблюдаемая концентрация HMP (3,4 пМ) приближается к медиане 2,8 пМ для ранее зарегистрированных концентраций (Carini et al., 2014; Suffridge et al., 2017, 2018). В текущем исследовании представлены первые образцы TRC, собранные во время фазы накопления весеннего цветения в Северной Атлантике (Behrenfeld et al., 2019). Мы предполагаем, что низкие наблюдаемые концентрации HMP являются результатом повышенного микробного спроса на этот ценный и широко необходимый TRC в период интенсивной микробной активности.
, 2014; Суффридж и др., 2017, 2018). Наша максимальная наблюдаемая концентрация HMP (3,4 пМ) приближается к медиане 2,8 пМ для ранее зарегистрированных концентраций (Carini et al., 2014; Suffridge et al., 2017, 2018). В текущем исследовании представлены первые образцы TRC, собранные во время фазы накопления весеннего цветения в Северной Атлантике (Behrenfeld et al., 2019). Мы предполагаем, что низкие наблюдаемые концентрации HMP являются результатом повышенного микробного спроса на этот ценный и широко необходимый TRC в период интенсивной микробной активности.
Распределение TRC по глубине имело схожие закономерности между станциями, несмотря на существенные различия в диапазонах концентраций (рис. 2, 3). Профили обычно были истощены вблизи поверхности по сравнению с максимумом на глубине 100 м (рис. 3). Мы наблюдали эту тенденцию на всех станциях, кроме станции 3 и станции 4-cHET, где наблюдалось поверхностное обогащение (рис. 3). Станции 3 и 4 были самыми северными станциями и единственными станциями, расположенными в субтропической океанографической области (Della Penna, Gaube, 2019) (рис. 1).Наблюдаемый среднеглубинный максимум часто совпадал с подповерхностными максимумами Chl a (рис. 1). Концентрация ТРК в целом снижалась ниже максимума ~100 м, а увеличения концентрации ТРК ниже 300 м не наблюдалось в двух глубинных пробах, взятых на станции 2.1 (1000 и 2600 м; таблица 2). Общая тенденция поверхностного истощения относительно максимума вблизи дна эвфотической зоны и низких концентраций на более глубоких глубинах аналогична ранее зарегистрированным тенденциям в других океанографических регионах для B1 и HMP, что указывает на потенциальную схему глобального распределения TRC (Sañudo-Wilhelmy и другие., 2012; Карини и др., 2014; Суффридж и др., 2017, 2018).
1).Наблюдаемый среднеглубинный максимум часто совпадал с подповерхностными максимумами Chl a (рис. 1). Концентрация ТРК в целом снижалась ниже максимума ~100 м, а увеличения концентрации ТРК ниже 300 м не наблюдалось в двух глубинных пробах, взятых на станции 2.1 (1000 и 2600 м; таблица 2). Общая тенденция поверхностного истощения относительно максимума вблизи дна эвфотической зоны и низких концентраций на более глубоких глубинах аналогична ранее зарегистрированным тенденциям в других океанографических регионах для B1 и HMP, что указывает на потенциальную схему глобального распределения TRC (Sañudo-Wilhelmy и другие., 2012; Карини и др., 2014; Суффридж и др., 2017, 2018).
Взаимосвязь между TRC и параметрами окружающей среды
Несмотря на сходство моделей распределения TRC на разрезе, отношения между распределением TRC и численностью бактерий, бактериальной продукцией, распределением Chl a, DOC и фосфатов не были универсальными для всех станций (дополнительный рисунок 1). Например, станции 1 и 2 имеют схожие характеристики TRC (рис. 3), но на станции 1 бактериальная численность, бактериальная продукция и Chl a положительно коррелируют с TRC, тогда как на станции 2 эти же параметры отрицательно коррелируют с TRC (рис. 5, Дополнительный рисунок 1).Точно так же TRC и фосфаты имеют отрицательную корреляцию на станции 1, но положительную корреляцию на станции 2. Величина обоих TRC и этих основных параметров была значительно выше на станции 2, чем на станции 1 (рис. 1), что позволяет предположить, что более высокая биологическая активность вызывает различные отношения между TRC и основными параметрами. На всех станциях, сходных со станцией 1 (например, станция 2.1, 3 и 4-cHET) или станция 2 (например, станция 3-HET и 3-HMP, станция 4 все, кроме cHET) (дополнительный рисунок 1).
Например, станции 1 и 2 имеют схожие характеристики TRC (рис. 3), но на станции 1 бактериальная численность, бактериальная продукция и Chl a положительно коррелируют с TRC, тогда как на станции 2 эти же параметры отрицательно коррелируют с TRC (рис. 5, Дополнительный рисунок 1).Точно так же TRC и фосфаты имеют отрицательную корреляцию на станции 1, но положительную корреляцию на станции 2. Величина обоих TRC и этих основных параметров была значительно выше на станции 2, чем на станции 1 (рис. 1), что позволяет предположить, что более высокая биологическая активность вызывает различные отношения между TRC и основными параметрами. На всех станциях, сходных со станцией 1 (например, станция 2.1, 3 и 4-cHET) или станция 2 (например, станция 3-HET и 3-HMP, станция 4 все, кроме cHET) (дополнительный рисунок 1).
Приведенные выше наблюдения показывают, что в масштабе разреза концентрации TRC не связаны с нашими измеренными биологическими и химическими параметрами ядра. Однако взаимосвязь между распределениями TRC и основными биологическими и химическими параметрами устойчива в локальном масштабе каждой отдельной станции. Иными словами, хотя характер распределения ТРК оставался сходным, их связь с биологическими и химическими параметрами различалась между станциями (рис. 1, 3, 5).Эти наблюдения позволили нам выдвинуть гипотезу о том, что различия в составе микробного сообщества между станциями ответственны за характер распределения ТРК и их взаимосвязь с биологическими и химическими параметрами.
Иными словами, хотя характер распределения ТРК оставался сходным, их связь с биологическими и химическими параметрами различалась между станциями (рис. 1, 3, 5).Эти наблюдения позволили нам выдвинуть гипотезу о том, что различия в составе микробного сообщества между станциями ответственны за характер распределения ТРК и их взаимосвязь с биологическими и химическими параметрами.
Взаимосвязь между TRC и составом микробного сообщества
Канонический анализ главных координат (CAP) показал, что микробные профили ASV относительно многочисленных таксонов, обнаруженных на каждой станции, различаются между станциями вблизи поверхности (5–75 м) и имеют некоторое перекрытие ниже эвфотической зоны (рис. 4).Мы исследовали взаимосвязь между 16S рРНК ASV (включая бактериопланктон, цианобактерии и эукариотические пластиды) на основе разнообразия микробного сообщества (Bolaños et al., 2020a), концентрациями TRC и параметрами окружающей среды. Все эти данные были собраны параллельно в ходе экспедиции NAAMES4, чтобы определить, создают ли различия в концентрациях TRC различия в микробном сообществе. Наш анализ показал, что микробные сообщества из верхних 100 м группируются близко к станции. Отдельные кластеры станций упорядочены в две большие группы по первому компоненту (станции 1, 2.1, 3 по сравнению со станциями 2,4). Эта кластеризация соответствовала корреляционным отношениям между TRC и биологическими и химическими параметрами, описанными выше (рис. 4). Эти наблюдения подтверждают интерпретацию того, что изменение состава планктонных микробных сообществ способствует наблюдаемым корреляциям между концентрациями TRC, концентрациями макроэлементов и биологическим статусом цветения.
Наш анализ показал, что микробные сообщества из верхних 100 м группируются близко к станции. Отдельные кластеры станций упорядочены в две большие группы по первому компоненту (станции 1, 2.1, 3 по сравнению со станциями 2,4). Эта кластеризация соответствовала корреляционным отношениям между TRC и биологическими и химическими параметрами, описанными выше (рис. 4). Эти наблюдения подтверждают интерпретацию того, что изменение состава планктонных микробных сообществ способствует наблюдаемым корреляциям между концентрациями TRC, концентрациями макроэлементов и биологическим статусом цветения.
Обнаружив доказательства связи между составом сообщества и концентрациями TRC, мы спросили, есть ли доказательства в данных генома, которые могут подтвердить связь метаболических стратегий B1 с концентрациями TRC.Мы идентифицировали ASV, которые были по-разному распространены в образцах с высокой и низкой концентрацией B1 (т. е. ASV, представляющие таксоны, которые могли бы объяснить отношения, наблюдаемые в анализе CAP). Концентрации TRC были разделены на «высокие» и «низкие» группы относительно средней концентрации TRC на каждой станции. Был проведен полный факторный анализ, анализирующий все комбинации TRC и ASV. Только B1 на станциях 1 и 2 демонстрируют различия в численности конкретных ASV (рис. 5). Анализ CAP также показал сильную дифференциацию между микробными сообществами на этих двух станциях.Таким образом, объединенные результаты этих двух анализов позволяют предположить, что концентрации B1 вносят важный вклад в наблюдаемые различия (рис. 4).
Концентрации TRC были разделены на «высокие» и «низкие» группы относительно средней концентрации TRC на каждой станции. Был проведен полный факторный анализ, анализирующий все комбинации TRC и ASV. Только B1 на станциях 1 и 2 демонстрируют различия в численности конкретных ASV (рис. 5). Анализ CAP также показал сильную дифференциацию между микробными сообществами на этих двух станциях.Таким образом, объединенные результаты этих двух анализов позволяют предположить, что концентрации B1 вносят важный вклад в наблюдаемые различия (рис. 4).
Станции 1 и 2 имеют противоположные отношения между TRC и микробным сообществом
На станции 1 олиготрофы, включая SAR11, были более многочисленны на глубине 25 и 75 м (где концентрации B1 были повышены), чем на глубине 5 м (где концентрации B1 были низкими) (рис. 5). И наоборот, на станции 2 копиотрофные флавобактерии были более многочисленны между поверхностью и 25 м (где концентрации B1 были низкими), чем на более глубоких глубинах (рис. 5).Стоит отметить, что самые низкие концентрации B1 на станции 2 примерно эквивалентны самым высоким концентрациям B1 на станции 1, а самые высокие концентрации B1 на станции 2 примерно в 5 раз превышают самые высокие концентрации B1 на станции 1 (рис. 5). На станции 1 градиент концентрации B1, по-видимому, не был связан со сложной реструктуризацией микробного сообщества, поскольку только несколько ASV реагировали на изменения концентрации B1 (рис. 5). Напротив, количество ответивших ASV на первых 25 м станции 2, их таксономическая принадлежность и высокие концентрации B1 на глубине 75 м и ниже показали, как наличие TRC может изменить сообщества (рис. 5).
5).Стоит отметить, что самые низкие концентрации B1 на станции 2 примерно эквивалентны самым высоким концентрациям B1 на станции 1, а самые высокие концентрации B1 на станции 2 примерно в 5 раз превышают самые высокие концентрации B1 на станции 1 (рис. 5). На станции 1 градиент концентрации B1, по-видимому, не был связан со сложной реструктуризацией микробного сообщества, поскольку только несколько ASV реагировали на изменения концентрации B1 (рис. 5). Напротив, количество ответивших ASV на первых 25 м станции 2, их таксономическая принадлежность и высокие концентрации B1 на глубине 75 м и ниже показали, как наличие TRC может изменить сообщества (рис. 5).
Станции 1 и 2 расположены в сходных физических условиях, что позволило нам провести поиск взаимосвязей между концентрациями ТРК и структурой микробного сообщества. Обе станции расположены примерно на одной широте и в одном океанографическом районе (рис. 1) (Behrenfeld et al., 2019). Глубина смешанных слоев была одинаковой на станциях 1 и 2 (80 и 65 м соответственно), однако, несмотря на подобные физические условия, наш многовариантный анализ и дифференциальный анализ разнообразия сообщества ASV показали потенциально контрастирующие стратегии сбора TRC на этих двух участках (рис. 4, 5). .
4, 5). .
Станция 1 была умеренно олиготрофной, с величинами ОКП, биологическими и химическими параметрами ниже, чем на станции 2. На станции 1 отношения между ОКП и другими измерениями были обратными таковым на станции 2. В частности, распределение ОКП положительно коррелировало с показателями роста и биомассы и отрицательно коррелировали с фосфатами (рис. 5), при этом интегрированный по глубине (0–150 м) уровень фосфатов был самым низким на разрезе (рис. 1). Тесная связь между концентрациями TRC и биологической активностью на станции 1 в сочетании со значениями продукции бактерий в 5 раз ниже, чем на станции 2, предполагает, что скорость производства и потребления микробами TRC связана со скоростью микробного метаболизма в олиготрофной среде.
Условия, обнаруженные на станции 2, соответствовали фазе накопления гипотезы восстановления возмущения; присутствовали самые высокие наблюдаемые концентрации TRC, продукция бактерий, Chl a и численность бактерий (Behrenfeld et al. , 2019). На этой станции TRC имели распределение, подобное питательным веществам, и отрицательно коррелировали с биомассой (рис. 5). Кроме того, концентрации фосфатов и TRC были низкими на поверхности и увеличивались с глубиной, в то время как другие биологические параметры были максимальными вблизи поверхности и уменьшались с глубиной.Фаза накопления весеннего цветения характеризуется периодом усиленного роста, как мы наблюдали (Behrenfeld, Boss, 2014, 2018). Одновременно с максимальной бактериальной продукцией мы наблюдали более низкие концентрации фосфатов и TRC в верхних 25 м и обилие ASV, соответствующих копиотрофным и быстрорастущим бактериальным линиям (рис. 5) (Kirchman, 2002; Pinhassi et al., 2004). Среди этих реагирующих бактерий наиболее представлены таксоны Flavobacteria. Это наблюдение может свидетельствовать о том, что поглощение B1 флавобактериями повлияло на вертикальную структуру концентраций B1, наблюдаемых в смешанном слое на этой станции.
, 2019). На этой станции TRC имели распределение, подобное питательным веществам, и отрицательно коррелировали с биомассой (рис. 5). Кроме того, концентрации фосфатов и TRC были низкими на поверхности и увеличивались с глубиной, в то время как другие биологические параметры были максимальными вблизи поверхности и уменьшались с глубиной.Фаза накопления весеннего цветения характеризуется периодом усиленного роста, как мы наблюдали (Behrenfeld, Boss, 2014, 2018). Одновременно с максимальной бактериальной продукцией мы наблюдали более низкие концентрации фосфатов и TRC в верхних 25 м и обилие ASV, соответствующих копиотрофным и быстрорастущим бактериальным линиям (рис. 5) (Kirchman, 2002; Pinhassi et al., 2004). Среди этих реагирующих бактерий наиболее представлены таксоны Flavobacteria. Это наблюдение может свидетельствовать о том, что поглощение B1 флавобактериями повлияло на вертикальную структуру концентраций B1, наблюдаемых в смешанном слое на этой станции.
Последующий сравнительный геномный анализ показал, что Flavobacteria ASV, которые были в большом количестве на 5 м и 25 м на станции 2, потенциально являются ауксотрофами B1, что подтверждает нашу гипотезу (рис. 5). Предыдущие исследования генома показали, что некоторые таксоны Flavobacteria являются ауксотрофами B1 и в некоторых случаях могут использовать энергию, полученную из протеородопсина, для получения витамина B1 из растворенного пула (Sañudo-Wilhelmy et al., 2014; Gómez-Consarnau et al., 2016, 2018). ). К сожалению, ауксотрофия B1 является парафилетической, а это означает, что данные ASV должны быть объединены с геномными данными для точного определения ауксотрофии витамина B1.Чтобы определить предполагаемый статус синтеза/ауксотрофии B1 дифференциально распространенных NAAMES flavobacteria ASV на станции 2, мы провели филогенетическую реконструкцию с использованием этих ASV, общедоступных SAG и последовательностей ASV flavobacteria из литературы (Alonso et al., 2007; Д’амброзио и др., 2014). Полученное дерево показало, что ASV NAAMES сгруппированы в три отдельные клады, и каждая из этих кладов включала по крайней мере один близкородственный SAG для сравнения генома (рис. 5).Филогенетическое размещение ASV флавобактерий NAAMES согласуется с предыдущими исследованиями, которые показали, что среди этих организмов существует высокая степень специализации ниш (Diez-Vives et al.
5). Предыдущие исследования генома показали, что некоторые таксоны Flavobacteria являются ауксотрофами B1 и в некоторых случаях могут использовать энергию, полученную из протеородопсина, для получения витамина B1 из растворенного пула (Sañudo-Wilhelmy et al., 2014; Gómez-Consarnau et al., 2016, 2018). ). К сожалению, ауксотрофия B1 является парафилетической, а это означает, что данные ASV должны быть объединены с геномными данными для точного определения ауксотрофии витамина B1.Чтобы определить предполагаемый статус синтеза/ауксотрофии B1 дифференциально распространенных NAAMES flavobacteria ASV на станции 2, мы провели филогенетическую реконструкцию с использованием этих ASV, общедоступных SAG и последовательностей ASV flavobacteria из литературы (Alonso et al., 2007; Д’амброзио и др., 2014). Полученное дерево показало, что ASV NAAMES сгруппированы в три отдельные клады, и каждая из этих кладов включала по крайней мере один близкородственный SAG для сравнения генома (рис. 5).Филогенетическое размещение ASV флавобактерий NAAMES согласуется с предыдущими исследованиями, которые показали, что среди этих организмов существует высокая степень специализации ниш (Diez-Vives et al. , 2019). Затем SAG опрашивали на предмет способности синтеза B1. Было определено, что SAG, которые кластеризуются ближе всего к ASV NAAMES, не имеют признаков способности к синтезу B1, в то время как доказательства способности к синтезу B1 были обнаружены в тех, которые были более филогенетически удалены (рис. 5).Эти результаты позволяют предположить, что NAAMES ASV являются предполагаемыми ауксотрофами витамина B1, способными истощать растворенный витамин B1 с глубины, где он был в изобилии на станции 2. Однако важно отметить, что SAG, где синтез B1 не был обнаружен, обычно имеют более низкую качество (полнота и загрязнение), чем те, в которых были обнаружены доказательства синтеза B1 (рис. 5). Хотя у этого анализа есть существенные ограничения (например, отсутствие доказательств не является свидетельством отсутствия), наши данные свидетельствуют о том, что поглощение витамина B1 ауксотрофными флавобактериями ответственно за истощение B1 на глубине 5–25 м по сравнению с 75 м на станции 2.
, 2019). Затем SAG опрашивали на предмет способности синтеза B1. Было определено, что SAG, которые кластеризуются ближе всего к ASV NAAMES, не имеют признаков способности к синтезу B1, в то время как доказательства способности к синтезу B1 были обнаружены в тех, которые были более филогенетически удалены (рис. 5).Эти результаты позволяют предположить, что NAAMES ASV являются предполагаемыми ауксотрофами витамина B1, способными истощать растворенный витамин B1 с глубины, где он был в изобилии на станции 2. Однако важно отметить, что SAG, где синтез B1 не был обнаружен, обычно имеют более низкую качество (полнота и загрязнение), чем те, в которых были обнаружены доказательства синтеза B1 (рис. 5). Хотя у этого анализа есть существенные ограничения (например, отсутствие доказательств не является свидетельством отсутствия), наши данные свидетельствуют о том, что поглощение витамина B1 ауксотрофными флавобактериями ответственно за истощение B1 на глубине 5–25 м по сравнению с 75 м на станции 2. .
.
Велоспорт TRC опосредует микробные взаимодействия: гипотетическая основа
Хотя из клеточной биологии и сравнительной геномики ясно, что TRC необходимы для роста многих планктонных таксонов, нет четкой основы для понимания того, как цикличность TRC влияет на структуру сообщества в океане. Данные, представленные в этом исследовании, представляют собой наиболее полный анализ океанографического распределения ТРК и его связи с микробным сообществом на сегодняшний день. Хотя мы считаем, что эта новая информация о циклах TRC является началом исследования важного и сложного процесса, на основе этих данных мы предлагаем гипотетическую основу, которая могла бы объяснить наблюдения этого исследования.Известно, что в условиях отсутствия цветения, когда чистая продуктивность сообщества планктона низка (на что указывает более низкая продукция бактерий, обилие бактерий и Хл а), структура микробного сообщества относительно стабильна, а разнообразие относительно велико (Fuhrman et al. , 2006). , 2015; Tai and Palenik, 2009; Teeling et al., 2012; Chow et al., 2013; Cram et al., 2015; Needham and Fuhrman, 2016). Мы предполагаем, что в этих условиях концентрации TRC приближаются к устойчивому состоянию, когда условия производства и потребления сбалансированы, а постоянный запас TRC положительно коррелирует с бактериальной продуктивностью, как видно на станции 1 (рис. 5).Напротив, стабильность сообщества нарушается во время цветения, когда микробное сообщество смещается в сторону доминирования одной группы или консорциума видов, что приводит к повышению микробной биомассы и метаболической активности (Teeling et al., 2012; Behrenfeld and Boss, 2018; Bolanos et al. и др., 2020b; Крамер и Графф, 2020; Ян и др., 2020). Мы предполагаем, что в условиях цветения, таких как те, которые наблюдаются на станции 2, условия производства и потребления ТОК становятся несбалансированными, и, в зависимости от статуса ауксотрофии ТОК цветущих организмов, спрос на ТОК может опережать производство и приводить к истощению ТОК (рис.
, 2006). , 2015; Tai and Palenik, 2009; Teeling et al., 2012; Chow et al., 2013; Cram et al., 2015; Needham and Fuhrman, 2016). Мы предполагаем, что в этих условиях концентрации TRC приближаются к устойчивому состоянию, когда условия производства и потребления сбалансированы, а постоянный запас TRC положительно коррелирует с бактериальной продуктивностью, как видно на станции 1 (рис. 5).Напротив, стабильность сообщества нарушается во время цветения, когда микробное сообщество смещается в сторону доминирования одной группы или консорциума видов, что приводит к повышению микробной биомассы и метаболической активности (Teeling et al., 2012; Behrenfeld and Boss, 2018; Bolanos et al. и др., 2020b; Крамер и Графф, 2020; Ян и др., 2020). Мы предполагаем, что в условиях цветения, таких как те, которые наблюдаются на станции 2, условия производства и потребления ТОК становятся несбалансированными, и, в зависимости от статуса ауксотрофии ТОК цветущих организмов, спрос на ТОК может опережать производство и приводить к истощению ТОК (рис. 5). .И наоборот, производство TRC может опережать спрос, что приводит к накоплению. Также вероятно, что по мере смены сообщества во время цветения (например, Teeling et al., 2012; Bolanos et al., 2020b) и замены одного доминирующего вида другим относительные темпы производства и потребления TRC могут измениться. Таким образом, мы предполагаем, что производство TRC опережало спрос до того, как мы занялись и взяли пробу на станции 2, что привело к избытку растворенных TRC, о чем свидетельствует увеличение концентрации TRC с глубиной на станции 2 (рис. 5).Впоследствии ауксотрофные флавобактерии TRC расцвели на глубине 5 и 25 м, что привело к опережающему росту спроса на TRC и, в конечном итоге, к истощению TRC на этих глубинах (рис. 5). Будущие исследования временных рядов потребуются для проверки этих гипотез, основанных на предположениях, но мы считаем, что вышеуказанная гипотеза согласуется с наблюдениями в один момент времени со станций 1 и 2.
5). .И наоборот, производство TRC может опережать спрос, что приводит к накоплению. Также вероятно, что по мере смены сообщества во время цветения (например, Teeling et al., 2012; Bolanos et al., 2020b) и замены одного доминирующего вида другим относительные темпы производства и потребления TRC могут измениться. Таким образом, мы предполагаем, что производство TRC опережало спрос до того, как мы занялись и взяли пробу на станции 2, что привело к избытку растворенных TRC, о чем свидетельствует увеличение концентрации TRC с глубиной на станции 2 (рис. 5).Впоследствии ауксотрофные флавобактерии TRC расцвели на глубине 5 и 25 м, что привело к опережающему росту спроса на TRC и, в конечном итоге, к истощению TRC на этих глубинах (рис. 5). Будущие исследования временных рядов потребуются для проверки этих гипотез, основанных на предположениях, но мы считаем, что вышеуказанная гипотеза согласуется с наблюдениями в один момент времени со станций 1 и 2.
Эти наблюдения за окружающей средой, показывающие дифференциальные отношения между микробными группами и наличием TRC, подтверждают результаты исследований клеточных культур и метагеномных исследований, которые демонстрируют, что смесь типов ауксотрофии TRC широко распространена в морских системах (Carini et al. , 2014; МакРоуз и др., 2014; Паэрл и др., 2016, 2018а,б; Гутовска и др., 2017). Состав морского микробного сообщества зависит от нескольких динамических во времени переменных, что делает рискованным использование дискретных во времени выборок для анализа влияния нескольких переменных. Тем не менее отношения, о которых мы сообщаем, поддерживают правдоподобную основу гипотез о влиянии TRC на сообщества.
, 2014; МакРоуз и др., 2014; Паэрл и др., 2016, 2018а,б; Гутовска и др., 2017). Состав морского микробного сообщества зависит от нескольких динамических во времени переменных, что делает рискованным использование дискретных во времени выборок для анализа влияния нескольких переменных. Тем не менее отношения, о которых мы сообщаем, поддерживают правдоподобную основу гипотез о влиянии TRC на сообщества.
Заявление о доступности данных
Полный набор данных TRC представлен в таблице 2. Другие наборы данных, представленные в этом исследовании, можно найти в онлайн-репозиториях.Названия репозиториев/репозиториев и номера доступа можно найти ниже: Биооптический архив и система хранения SeaWiFS (SeaBASS, seabass.gsfc.nasa.gov/Investigator/Giovannoni,%20Stephen) в качестве связанных файлов, NCBI BioProject — PRJNA627189 (регистрационные номера от SRR11596939 до SRR11596981).
Вклад авторов
Проект был запланирован CS, SG, MB, AW и LB. Образцы были собраны LB и MB. Анализ проб был проведен CS, LB и JM. Анализ данных был проведен CS, LB, JM и KB.Рукопись была написана CS и отредактирована LB, KB, AW, JM, MB и SG. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Анализ проб был проведен CS, LB и JM. Анализ данных был проведен CS, LB, JM и KB.Рукопись была написана CS и отредактирована LB, KB, AW, JM, MB и SG. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа финансировалась за счет гранта Национального научного фонда «Размеры совместных исследований»: «Разгадка сложности циклов тиамина и воздействия на микробные сети» для SG (DEB-1639033) и AW (DEB-1638928). Поддержка операций корабля, сбора проб и анализа основных параметров была предоставлена грантом программы NASA NAAMES для SG (NNX15AE70G) и MB (NNX15AE30G).Приборы для масс-спектрометрии были поддержаны грантом NIH 1S10RR022589-01.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Элизабет Бреннан, Стивена Ноэля, Сару Вульф и Веронику Кивенсон за их полезные комментарии во время написания и редактирования этой рукописи.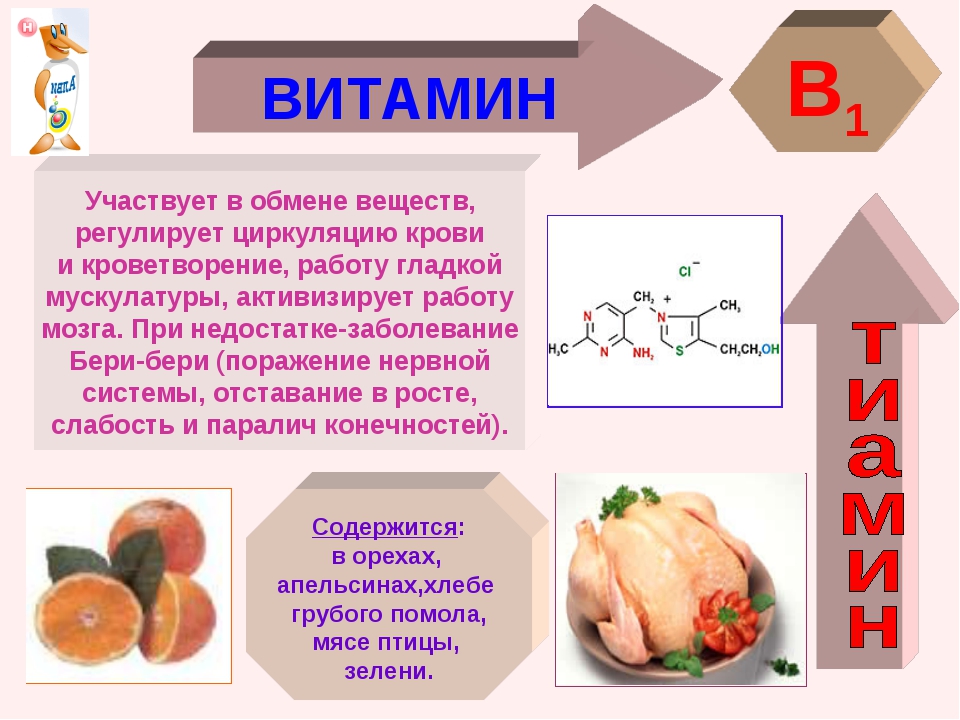 Мы благодарим Марка Дасенко и Орегонского государственного университета CGRB за подготовку библиотеки ампликонов и секвенирование.Мы благодарим капитана и экипаж НИС «Атлантис» за помощь в сборе проб. Мы благодарим сообщество NAAMES за поддержку.
Мы благодарим Марка Дасенко и Орегонского государственного университета CGRB за подготовку библиотеки ампликонов и секвенирование.Мы благодарим капитана и экипаж НИС «Атлантис» за помощь в сборе проб. Мы благодарим сообщество NAAMES за поддержку.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmars.2020.606342/full#supplementary-material
.Дополнительный рисунок 1. Корреляции Спирмена между концентрациями TRC и основными биологическими, химическими и физическими параметрами на каждой станции.Для этих параметров были рассчитаны корреляционные матрицы Спирмена на глубинах, где были отобраны пробы TRC. Сила и величина корреляции (ро Спирмена) изображаются с помощью цвета. Статистическая значимость не оценивалась из-за малого количества выборок. Параметры были упорядочены с использованием иерархической кластеризации.
Дополнительный рисунок 2. Филогенетический контекст флавобактерий NAAMES предполагает ауксотрофию B1. Это полностью аннотированная версия рисунка 5. Филогенетические отношения последовательностей гена 16S рРНК, извлеченных из экспедиции NAAMES4 в изобилии на глубине 5–25 м на станции 2, отмечены красным, и эталонных последовательностей из прибрежных районов Северного моря (Alonso et al., 2007) и прибрежные районы Северной Каролины (D’ambrosio et al., 2014), отмеченные черным цветом. Отдельные амплифицированные геномы выделены зеленым цветом. Отдельные амплифицированные геномы, кодирующие thiC (предполагаемые ауксотрофы витамина B1), обозначены звездочкой (дополнительная таблица 3). Филогенетическое дерево было рассчитано с использованием программного инструмента MAFFT, Method Neighbor-Joining, Jukes-Cantor Model, Bootstrap resampling 100. Полноразмерные последовательности были обрезаны до NAAMES ASV (330 п.н.).
Филогенетический контекст флавобактерий NAAMES предполагает ауксотрофию B1. Это полностью аннотированная версия рисунка 5. Филогенетические отношения последовательностей гена 16S рРНК, извлеченных из экспедиции NAAMES4 в изобилии на глубине 5–25 м на станции 2, отмечены красным, и эталонных последовательностей из прибрежных районов Северного моря (Alonso et al., 2007) и прибрежные районы Северной Каролины (D’ambrosio et al., 2014), отмеченные черным цветом. Отдельные амплифицированные геномы выделены зеленым цветом. Отдельные амплифицированные геномы, кодирующие thiC (предполагаемые ауксотрофы витамина B1), обозначены звездочкой (дополнительная таблица 3). Филогенетическое дерево было рассчитано с использованием программного инструмента MAFFT, Method Neighbor-Joining, Jukes-Cantor Model, Bootstrap resampling 100. Полноразмерные последовательности были обрезаны до NAAMES ASV (330 п.н.).
Дополнительная таблица 1. Значительная дифференциальная численность ASV при низких и высоких концентрациях TRC на каждой станции. ASV, лежащие в основе статистики HEAT, представлены вместе с полным таксономическим присвоением. Порог P с поправкой <0,01 был установлен для определения Log2foldchange > |2| как существенное изменение.
ASV, лежащие в основе статистики HEAT, представлены вместе с полным таксономическим присвоением. Порог P с поправкой <0,01 был установлен для определения Log2foldchange > |2| как существенное изменение.
Дополнительная таблица 2. Общедоступные амплифицированные геномы Flavobacteria, полученные из базы данных IMG, которые были исследованы на предмет биосинтеза и транспортных белков, связанных с TRC.
Дополнительная таблица 3. Flavobacteria SAG Полнота генома.
Дополнительная таблица 4. Ключевые ферменты для пути de novo синтеза витамина B1, присутствие которого в SAG флавобактерий определено с помощью анализа HMMER.
Каталожные номера
Алонсо, К., Варнеке, Ф., Аманн, Р., и Пернталер, Дж. (2007). Высокое локальное и глобальное разнообразие флавобактерий в морском планктоне. Окружающая среда. микробиол. 9, 1253–1266. doi: 10.1111/j.1462-2920.2007.01244.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Атильо, Р. М., Арахчилаге Г.М., Гринли Э.Б., Кнехт К.М. и Брейкер Р.Р. (2019). Класс бактериальных рибопереключателей для предшественника тиамина HMP-PP использует аптамер, встроенный в терминатор. Элиф 8:e45210. doi: 10.7554/eLife.45210.020
М., Арахчилаге Г.М., Гринли Э.Б., Кнехт К.М. и Брейкер Р.Р. (2019). Класс бактериальных рибопереключателей для предшественника тиамина HMP-PP использует аптамер, встроенный в терминатор. Элиф 8:e45210. doi: 10.7554/eLife.45210.020
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бетге, Н., Графф, Дж. Р., Беренфельд, М. Дж., и Карлсон, К. А. (2020). Чистая продукция сообщества, накопление растворенного органического углерода и вертикальный экспорт в западной части Северной Атлантики. Перед. Морские науки. 7:227. doi: 10.3389/fmars.2020.00227
Полнотекстовая перекрестная ссылка | Академия Google
Begley, T.P., Downs, D.M., Ealick, S.E., Mclafferty, F.W., van Loon, A., Taylor, S., et al. (1999). Биосинтез тиамина у прокариот. Арх. микробиол. 171, 293–300. дои: 10.1007/s002030050713
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Беренфельд, М.Дж., и Босс, Э.С. (2014). Возрождение экологических основ цветения океанического планктона. Энн. Преподобный Мар. 6, 167–194. doi: 10.1146/annurev-marine-052913-021325
Возрождение экологических основ цветения океанического планктона. Энн. Преподобный Мар. 6, 167–194. doi: 10.1146/annurev-marine-052913-021325
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Беренфельд, М.Дж., и Босс, Э.С. (2018). Учебное пособие для студентов по гипотезам цветения в контексте годовых циклов фитопланктона. Глоб. Чанг. биол. 24, 55–77. doi: 10.1111/gcb.13858
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Беренфельд, М. Дж., Мур, Р. Х., Хостетлер, К.A., Graff, J., Gaube, P., Russell, L.M., et al. (2019). Исследование аэрозолей и морских экосистем Северной Атлантики (NAAMES): научный мотив и обзор миссии. Перед. Морские науки. 6:122. doi: 10.3389/fmars.2019.00122
Полнотекстовая перекрестная ссылка | Академия Google
Боланьос, Л. М., Чой, С. Дж., Уорден, А. З., Беате, Н., Карлсон, К. А., и Джованнони, С. Дж. (2020a). Сезонность состава микробного сообщества Северной Атлантики. Перед. Морские науки. (на рассмотрении).
Перед. Морские науки. (на рассмотрении).
Боланьос, Л. М., Карп-Босс, Л., Чой, С. Дж., Уорден, А. З., Графф, Дж. Р., Хентьенс, Н., и др. (2020б). Мелкий фитопланктон доминирует в биомассе западной части Северной Атлантики. ISME J. 14, 1663–1674. doi: 10.1038/s41396-020-0636-0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Каллахан, Б. Дж., Макмерди, П. Дж., Розен, М. Дж., Хан, А. В., Джонсон, А. Дж., и Холмс, С. П. (2016). DADA2: вывод образца с высоким разрешением на основе данных ампликона Illumina. Нац. Методы 13, 581–583. doi: 10.1038/nmeth.3869
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Carini, P., Campbel, E.O., Morre, J., Sanudo-Wilhelmy, S.A., Thrash, J.C., Bennett, S.E., et al. (2014). Обнаружение потребности роста SAR11 в предшественнике пиримидина тиамина и его распространение в Саргассовом море. ISME J. 8, 1727–1738. doi: 10.1038/ismej.2014. 61
61
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Карлуччи, А.Ф. (1970). Экология планктона у побережья Ла-Хойя, Калифорния, в период с апреля по сентябрь 1967 г. Часть 2. Витамин В, тиамин и биотин. Бык. Scripps Instn Oceanogr. Тех. сер. 17, 23–32.
Академия Google
Карлуччи, А.Ф., и Боуз, П.М. (1972). Определение витамина В12, тиамина и биотина в водах озера Тахо с использованием модифицированных методов морского биоанализа. Лимнол. океаногр. 17, 774–777. doi: 10.4319/lo.1972.17.5.0774
Полнотекстовая перекрестная ссылка | Академия Google
Карлуччи, А.Ф., Зильбернагель С.Б. и Макнелли П.М. (1969). Влияние температуры и солнечной радиации на стойкость витамина В12, тиамина и биотина в морской воде. J. Phycol. 5, 302–305. doi: 10.1111/j.1529-8817.1969.tb02618.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чаттерджи, А., Юргенсон, К.Т. , Шредер, Ф.К., Илик, С.Е., и Бегли, Т.П. (2006). Биосинтез тиамина у эукариот: характеристика связанного с ферментом продукта тиазолсинтазы из Saccharomyces c erevisiae и его значение в биосинтезе тиазола. Дж. Ам. хим. соц. 128, 7158–7159. дои: 10.1021/ja061413o
, Шредер, Ф.К., Илик, С.Е., и Бегли, Т.П. (2006). Биосинтез тиамина у эукариот: характеристика связанного с ферментом продукта тиазолсинтазы из Saccharomyces c erevisiae и его значение в биосинтезе тиазола. Дж. Ам. хим. соц. 128, 7158–7159. дои: 10.1021/ja061413o
Полнотекстовая перекрестная ссылка | Академия Google
Choi, C.J., Bachy, C., Jaeger, G.S., Poirier, C., Sudek, L., Sarma, V., et al. (2017). Недавно обнаруженные линии морских пластид с глубоким разветвлением численно редки, но распространены по всему миру. Курс. биол. 27, Р15–Р16. doi: 10.1016/j.cub.2016.11.032
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чоу, К.Э.Т., Сачдева, Р., Cram, J.A., Steele, J.A., Needham, D.M., Patel, A., et al. (2013). Временная изменчивость и когерентность бактериальных сообществ эвфотической зоны за десятилетие в Южно-Калифорнийском заливе. ISME J. 7, 2259–2273. doi: 10.1038/ismej.2013.122
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Cram, J. A., Chow, C.E., Sachdeva, R., Needham, D.M., Parada, A.E., Steele, J.A., et al. (2015). Сезонная и межгодовая изменчивость сообщества морского бактериопланктона по толще воды за десять лет. ИСМЭ J . 9, 563–580. doi: 10.1038/ismej.2014.153
A., Chow, C.E., Sachdeva, R., Needham, D.M., Parada, A.E., Steele, J.A., et al. (2015). Сезонная и межгодовая изменчивость сообщества морского бактериопланктона по толще воды за десять лет. ИСМЭ J . 9, 563–580. doi: 10.1038/ismej.2014.153
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Крофт, М.Т., Мулен, М., Уэбб, М.Е., и Смит, А.Г. (2007). Биосинтез тиамина у водорослей регулируется рибопереключателями. Проц. Натл. акад. науч. США 104, 20770–20775. doi: 10.1073/pnas.0705786105
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Д’амброзио Л., Зиерфогель К., Макгрегор Б., Теске А.и Арности, К. (2014). Состав и ферментативная функция ассоциированных с частицами и свободноживущих бактерий: сравнение прибрежных и морских. ISME J. 8, 2167–2179. doi: 10.1038/ismej.2014.67
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дэвис, Н. М., Проктор, Д. М. , Холмс, С. П., Релман, Д. А., и Каллахан, Б. Дж. (2018). Простая статистическая идентификация и удаление загрязняющих последовательностей в данных маркерного гена и метагеномики. Микробиом 6:226.doi: 10.1186/s40168-018-0605-2
, Холмс, С. П., Релман, Д. А., и Каллахан, Б. Дж. (2018). Простая статистическая идентификация и удаление загрязняющих последовательностей в данных маркерного гена и метагеномики. Микробиом 6:226.doi: 10.1186/s40168-018-0605-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Делла Пенна, А., и Гобе, П. (2019). Обзор (суб) мезомасштабной динамики океана для полевой программы NAAMES. Перед. Морские науки. 6:384. doi: 10.3389/fmars.2019.00384
Полнотекстовая перекрестная ссылка | Академия Google
Diez-Vives, C., Nielsen, S., Sanchez, P., Palenzuela, O., Ferrera, I., Sebastian, M., et al. (2019). Выделение экологически обособленных единиц морских Bacteroidetes в северо-западной части Средиземного моря. Мол. Экол. 28, 2846–2859. doi: 10.1111/mec.15068
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Донован, П.Д., Холланд, Л.М., Ломбарди, Л., Кофлан, А.Ю., Хиггинс, Д. Г., Вулф, К.Х., и соавт. (2018). Зависимая от рибопереключателя TPP регуляция древнего переносчика тиамина у Candida. ПЛОС Жене . 14:e1007429. doi: 10.1371/journal.pgen.1007429
Г., Вулф, К.Х., и соавт. (2018). Зависимая от рибопереключателя TPP регуляция древнего переносчика тиамина у Candida. ПЛОС Жене . 14:e1007429. doi: 10.1371/journal.pgen.1007429
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фокс, Дж., Behrenfeld, M.J., Haëntjens, N., Chase, A., Kramer, S.J., Boss, E., et al. (2020). Рост и продуктивность фитопланктона в западной части Северной Атлантики: наблюдения за региональной изменчивостью в ходе полевых кампаний NAAMES. Перед. Морские науки. 7:24. doi: 10.3389/fmars.2020.00024
Полнотекстовая перекрестная ссылка | Академия Google
Фурман, Дж. А., Крам, Дж. А., и Нидхэм, Д. М. (2015). Динамика морских микробных сообществ и их экологическая интерпретация. Нац.Преподобный Микробиолог. 13, 133–146. doi: 10.1038/nrmicro3417
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фурман, Дж. А., Хьюсон, И., Швальбах, М. С., Стил, Дж. А., Браун, М. В., и Наим, С. (2006). Ежегодно повторяющиеся бактериальные сообщества можно предсказать, исходя из условий океана. Проц. Натл. акад. науч. США 103, 13104–13109. doi: 10.1073/pnas.0602399103
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джованнони, С.Дж., Раппе, М.С., Вергин, К.Л., и Адэр, Н.Л. (1996). Гены 16S рРНК выявляют стратифицированные популяции бактериопланктона открытого океана, связанные с зелеными несерными бактериями. Проц. Натл. акад. науч. США 93, 7979–7984. doi: 10.1073/pnas.93.15.7979
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gold, K., Roels, O.A., and Bank, H. (1966). Температурно-зависимая деструкция тиамина в морской воде. Лимнол. океаногр. 11, 410–413. дои: 10.4319/ло.1966.11.3.0410
Полнотекстовая перекрестная ссылка | Академия Google
Гомес-Консарнау, Л., Гонсалес, Х.М., Ридель, Т., Йенике, С., Вагнер-Доблер, И., Санудо-Вильгельми, С. А., и др. (2016). Усиленный светом протеородопсин рост связан с приобретением витамина B морскими флавобактериями. ISME J. 10, 1102–1112. doi: 10.1038/ismej.2015.196
А., и др. (2016). Усиленный светом протеородопсин рост связан с приобретением витамина B морскими флавобактериями. ISME J. 10, 1102–1112. doi: 10.1038/ismej.2015.196
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гомес-Консарнау, Л., Сачдева, Р., Гиффорд, С.М., Каттер Л.С., Фурман Дж.А., Санудо-Вильгельми С.А. и соавт. (2018). Мозаика синтеза и использования витамина В в природном морском микробном сообществе. Окружающая среда. микробиол. 20, 2809–2823. дои: 10.1111/1462-2920.14133
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гутовска М.А., Шоме Б., Судек С., МакРоуз Д.Л., Гамильтон М., Джованнони С.Дж. и др. (2017). Глобально важные гаптофитные водоросли используют экзогенные пиримидиновые соединения более эффективно, чем тиамин. MBio 8, e01459–e01417. doi: 10.1128/mBio.01459-17
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Heal, K. R., Carlson, L.T., Devol, A.H., Armbrust, E., Moffett, J.W., Stahl, D.A., et al. (2014). Определение четырех форм витамина В12 и других витаминов группы В в морской воде методом жидкостной хроматографии/тандемной масс-спектрометрии. Быстрая общ. Масс-спектрометрия 28, 2398–2404. doi: 10.1002/rcm.7040
R., Carlson, L.T., Devol, A.H., Armbrust, E., Moffett, J.W., Stahl, D.A., et al. (2014). Определение четырех форм витамина В12 и других витаминов группы В в морской воде методом жидкостной хроматографии/тандемной масс-спектрометрии. Быстрая общ. Масс-спектрометрия 28, 2398–2404. doi: 10.1002/rcm.7040
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хантер, С.и Провасоли, Л. (1964). Питание водорослей. Анну. Преподобный Завод Физиол. 15, 37–56. doi: 10.1146/annurev.pp.15.060164.000345
Полнотекстовая перекрестная ссылка | Академия Google
Jurgenson, C.T., Begley, T.P., and Ealick, S.E. (2009). Структурно-биохимические основы биосинтеза тиамина. Анну. Преподобный Биохим. 78, 569–603. doi: 10.1146/annurev.biochem.78.072407.102340
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кох, Ф., Хаттен-Леманн, Т.К., Голески, Дж.А., Санудо-Вильгельми, С.А., Фишер, Н.С., и Гоблер, К. Дж. (2012). Поглощение и круговорот витаминов B1 и B12 сообществами планктона в прибрежных экосистемах. Перед. микробиол. 3:363. doi: 10.3389/fmicb.2012.00363
Дж. (2012). Поглощение и круговорот витаминов B1 и B12 сообществами планктона в прибрежных экосистемах. Перед. микробиол. 3:363. doi: 10.3389/fmicb.2012.00363
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кох Ф., Санудо-Вильгельми С. А., Фишер Н. С. и Гоблер С. Дж. (2013). Влияние витаминов В1 и В12 на динамику цветения вредоносной бурой водоросли. Лимнол.океаногр. 58, 1761–1774. doi: 10.4319/lo.2013.58.5.1761
Полнотекстовая перекрестная ссылка | Академия Google
Крамер, С.Дж., и Графф, Дж.Р. (2020). Состав сообщества фитопланктона, определенный на основе совместной изменчивости пигментов фитопланктона в ходе полевой кампании NAAMES. Перед. Морские науки. 7:215. doi: 10.3389/fmars.2020.00215
Полнотекстовая перекрестная ссылка | Академия Google
Ландри З., Свон Б.К., Херндл Г.Дж., Степанаускас Р. и Джованнони С.Дж. (2017). Геномы SAR202 из темного океана предсказывают пути окисления неподатливого растворенного органического вещества. MBio 8:e00413-17. doi: 10.1128/mBio.00413-17
MBio 8:e00413-17. doi: 10.1128/mBio.00413-17
Полнотекстовая перекрестная ссылка | Академия Google
Лав, М. И., Хубер, В., и Андерс, С. (2014). Модерированная оценка изменения кратности и дисперсии для данных секвенирования РНК с помощью DESeq2. Геном Биол. 15:550. doi: 10.1186/s13059-014-0550-8
Полнотекстовая перекрестная ссылка | Академия Google
Мартин, П., Лэмпитт, Р. С., Джейн Перри, М., Сандерс, Р., Ли, К., и Д’асаро, Э. (2011). Экспорт и поток мезопелагических частиц во время весеннего цветения диатомовых водорослей в Северной Атлантике. Deep Sea Res. Часть I Океаногр. Рез. Пап. 58, 338–349. doi: 10.1016/j.dsr.2011.01.006
Полнотекстовая перекрестная ссылка | Академия Google
Мопен-Ферлоу, Дж. А. (2018). Метаболизм и регуляция витамина B1 (тиамина) у архей. Витамины группы В — текущее использование и перспективы , 9–31. doi: 10.5772/intechopen.77170
Полнотекстовая перекрестная ссылка | Академия Google
Макмерди, П. Дж., и Холмс, С. (2013). phyloseq: пакет R для воспроизводимого интерактивного анализа и графики данных переписи микробиома. PLoS ONE 8:e61217. doi: 10.1371/journal.pone.0061217
Дж., и Холмс, С. (2013). phyloseq: пакет R для воспроизводимого интерактивного анализа и графики данных переписи микробиома. PLoS ONE 8:e61217. doi: 10.1371/journal.pone.0061217
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
McRose, D., Guo, J., Monier, A., Sudek, S., Wilken, S., Yan, S., et al. (2014). Альтернативы поглощению витамина B1 были обнаружены с открытием рибопереключателей во многих линиях морских эукариот. ISME J. 8, 2517–2529. doi: 10.1038/ismej.2014.146
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мохика, К.Д.А., Карлсон, К.А., и Беренфельд, М.Дж. (2020). Регуляция флуоресцентных гетеротрофных субпопуляций прокариот с низким и высоким содержанием нуклеиновых кислот и связь с вирусной смертностью в естественных сообществах прокариотических вирусов. Микроб. Экол. 79, 213–230. doi: 10.1007/s00248-019-01393-9
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Монтеверде, Д. Р., Гомес-Консарнау, Л., Каттер, Л., Чонг, Л., Берельсон, В., и Санудо-Вильгельми, С.А. (2015). Витамин B1 в морских отложениях: градиент концентрации воды в порах вызывает движение бентоса с потенциальными биологическими последствиями. Перед. микробиол. 6:434. doi: 10.3389/fmicb.2015.00434
Р., Гомес-Консарнау, Л., Каттер, Л., Чонг, Л., Берельсон, В., и Санудо-Вильгельми, С.А. (2015). Витамин B1 в морских отложениях: градиент концентрации воды в порах вызывает движение бентоса с потенциальными биологическими последствиями. Перед. микробиол. 6:434. doi: 10.3389/fmicb.2015.00434
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Монтеверде, Д. Р., Гомес-Консарнау, Л., Суффридж, К., и Санудо-Вильгельми, С. А. (2017). Жизненное использование витаминов группы В на ранней Земле. Геобиология 15, 3–18. doi: 10.1111/gbi.12202
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Моррис, Р. М., Вергин, К. Л., Чо, Дж.-К., Раппе, М. С., Карлсон, К. А., и Джованнони, С. Дж. (2005). Временная и пространственная реакция линий бактериопланктона на ежегодный конвективный переворот на участке изучения временных рядов Бермудских островов в Атлантике. Лимнол. океаногр. 50, 1687–1696. doi: 10.4319/lo. 2005.50.5.1687
2005.50.5.1687
Полнотекстовая перекрестная ссылка | Академия Google
Нидхэм, Д.М. и Фурман, Дж. А. (2016). Ярко выраженная суточная смена фитопланктона, архей и бактерий после весеннего цветения. Нац. микробиол. 1:16005. doi: 10.1038/nmicrobiol.2016.5
Реферат PubMed | Полнотекстовая перекрестная ссылка
Okbamichael, M., and Sanudo-Wilhelmy, S.A. (2005). Прямое определение витамина B-1 в морской воде методом твердофазной экстракции и количественного определения с помощью высокоэффективной жидкостной хроматографии. Лимнол. океаногр. Методы 3, 241–246. дои: 10.4319/лом.2005.3.241
Полнотекстовая перекрестная ссылка | Академия Google
Оксанен Дж., Киндт Р., Лежандр П., О’Хара Б., Стивенс М.Х.Х., Оксанен М.Дж. и др. (2007). Веганский пакет. Комм. Экол. Пакет 10, 631–637. Доступно в Интернете по адресу: https://CRAN.R-project.org/package=vegan
.Академия Google
Паерл, Р. В., Бертран, Э. М., Аллен, А. Э., Паленик, Б., и Азам, Ф. (2015). Экофизиология витамина B1 морских пикоэукариотических водорослей: штаммоспецифические различия и новая роль бактерий в круговороте витаминов. Лимнол. океаногр. 60, 215–228. doi: 10.1002/lno.10009
М., Аллен, А. Э., Паленик, Б., и Азам, Ф. (2015). Экофизиология витамина B1 морских пикоэукариотических водорослей: штаммоспецифические различия и новая роль бактерий в круговороте витаминов. Лимнол. океаногр. 60, 215–228. doi: 10.1002/lno.10009
Полнотекстовая перекрестная ссылка | Академия Google
Paerl, R.W., Bertrand, E.M., Rowland, E., Schatt, P., Mehiri, M., Niehaus, T.D., et al. (2018а). Карбокситиазол является ключевой микробной питательной валютой и важным компонентом биосинтеза тиамина. науч. Респ. 8:5940. doi: 10.1038/s41598-018-27042-8
Полнотекстовая перекрестная ссылка | Академия Google
Паерл, Р. В., Буже, Ф.-Ю., Лозано, Дж.-C., Verge, V., Schatt, P., Allen, E.E., et al. (2016). Использование предшественников витамина B1, полученных из планктона, особенно предшественника, связанного с тиазолом, ключевым морским пикоэукариотическим фитопланктоном. ISME J. 11, 753–765. doi: 10.1038/ismej.2016.145
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Paerl, R. W., Sundh, J., Tan, D., Svenningsen, S.L., Hylander, S., Pinhassi, J., et al. (2018б). Преобладающая зависимость бактериопланктона от экзогенного витамина B1 и наличия предшественников. Проц. Натл. акад. науч. США 115, E10447–E10456. doi: 10.1073/pnas.1806425115
W., Sundh, J., Tan, D., Svenningsen, S.L., Hylander, S., Pinhassi, J., et al. (2018б). Преобладающая зависимость бактериопланктона от экзогенного витамина B1 и наличия предшественников. Проц. Натл. акад. науч. США 115, E10447–E10456. doi: 10.1073/pnas.1806425115
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пинхасси, Дж., Сала, М.М., Хавскум, Х., Петерс, Ф., Гуадайол, О., Малитс, А., и др. (2004). Изменения состава бактериопланктона при различных режимах фитопланктона. Заяв. Окружающая среда. микробиол. 70, 6753–6766. doi: 10.1128/AEM.70.11.6753-6766.2004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Провасоли, Л.и Карлуччи, А.Ф. (1974). Витамины и регуляторы роста. Food Agric.Organ. UA 10, 741–787.
Академия Google
R Основная команда. (2015). R: Язык и среда для статистических вычислений . Вена: Фонд статистических вычислений R.
Академия Google
Рапала-Козик, М. (2011). «Витамин B1 (тиамин): кофактор для ферментов, участвующих в основных метаболических путях, и средство защиты от стресса окружающей среды», в Advances in Botanical Research , eds F.Ребей и Р. Дус (Лондон: Elsevier), 37–91. дои: 10.1016/B978-0-12-386479-6.00004-4
(2011). «Витамин B1 (тиамин): кофактор для ферментов, участвующих в основных метаболических путях, и средство защиты от стресса окружающей среды», в Advances in Botanical Research , eds F.Ребей и Р. Дус (Лондон: Elsevier), 37–91. дои: 10.1016/B978-0-12-386479-6.00004-4
Полнотекстовая перекрестная ссылка | Академия Google
Саньюдо-Вильгельми, С.А., Каттер, Л.С., Дурасо, Р., Смайл, Э.А., Гомес-Консарнау, Л., Уэбб, Э.А., и соавт. (2012). Множественное истощение витаминов группы В на больших участках побережья океана. Проц. Натл. акад. науч. США 109, 14041–14045. doi: 10.1073/pnas.1208755109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Саньюдо-Вильгельми, С.А., Гомес-Консарнау, Л., Суффридж, К., и Уэбб, Э.А. (2014). Роль витаминов группы В в морской биогеохимии. Энн. Преподобный Мар. 6, 339–367. doi: 10.1146/annurev-marine-120710-100912
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Шоуэн, Р. (1998). «Тиаминзависимые ферменты», в Комплексный биологический катализ , изд. SM (Сан-Диего: Academic Press), 212–266.
(1998). «Тиаминзависимые ферменты», в Комплексный биологический катализ , изд. SM (Сан-Диего: Academic Press), 212–266.
Судек, С., Эверроуд, Р. К., Гехман, А. Л. М., Смит, Дж.М., Пуарье С.Л., Чавес Ф.П. и соавт. (2015). Распределение цианобактерий по физико-химическому градиенту в северо-восточной части Тихого океана. Окружающая среда. микробиол. 17, 3692–3707. дои: 10.1111/1462-2920.12742
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Саффридж, К., Каттер, Л., и Санудо-Вильгельми, С. (2017). Новый аналитический метод для прямого измерения взвешенных и растворенных витаминов группы В и их родственных соединений в морской воде. Перед.Морские науки. 4:11. doi: 10.3389/fmars.2017.00011
Полнотекстовая перекрестная ссылка | Академия Google
Suffridge, C.P., Gómez-Consarnau, L., Monteverde, D.R., Cutter, L., Aristegui, J., Alvarez-Salgado, X.A., et al. (2018). Витамины группы В и их аналоги как потенциальные факторы состава микробного сообщества в олиготрофной морской экосистеме. Ж. Геофиз. Рез. Биогеология. 123, 2890–2907. дои: 10.1029/2018JG004554
Ж. Геофиз. Рез. Биогеология. 123, 2890–2907. дои: 10.1029/2018JG004554
Полнотекстовая перекрестная ссылка | Академия Google
Тан, Ю.З., Кох Ф. и Гоблер С.Дж. (2010). Наиболее вредными видами цветения водорослей являются ауксотрофы витаминов B1 и B12. Проц. Натл. акад. науч. США 107, 20756–20761. doi: 10.1073/pnas.1009566107
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Teeling, H., Fuchs, B.M., Becher, D., Klockow, C., Gardebrecht, A., Bennke, C.M., et al. (2012). Контролируемая субстратом последовательность популяций морского бактериопланктона, вызванная цветением фитопланктона. Наука 336, 608–611.doi: 10.1126/science.1218344
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Vergin, K.L., Beszteri, B., Monier, A., Thrash, J.C., Temperton, B., Treusch, A.H., et al. (2013). Динамика экотипа SAR11 с высоким разрешением на участке исследования временных рядов Бермудских островов в Атлантическом океане путем филогенетического размещения пиросеквенций. ISME J. 7, 1322–1332. doi: 10.1038/ismej.2013.32
ISME J. 7, 1322–1332. doi: 10.1038/ismej.2013.32
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уикхем, Х.(2009). Ggplot2: элегантная графика для анализа данных. Дордрехт, Нью-Йорк, штат Нью-Йорк: Springer. дои: 10.1007/978-0-387-98141-3
Полнотекстовая перекрестная ссылка | Академия Google
Ян Б., Босс Э. С., Хантьенс Н., Лонг М. С., Беренфельд М. Дж., Эвелет Р. и др. (2020). Фенология фитопланктона в Северной Атлантике: результаты профилирования поплавковых измерений. Перед. Морские науки. 7:139. doi: 10.3389/fmars.2020.00139
Полнотекстовая перекрестная ссылка | Академия Google
Новостная статья: Смертельный дефицит в основе загадки окружающей среды
Исследователи ломают голову над широко распространенной нехваткой витамина B, которая, по-видимому, убивает диких животных .
Весной и летом на скалистых побережьях Балтийского моря обычно можно увидеть многолюдные колонии уток, называемых гага обыкновенной ( Somateria mollissima ), и других диких птиц, гнездящихся на скалистых побережьях. Тысячи нетерпеливых молодых родителей соревнуются за лучшие места для строительства гнезд и добычи пищи для своих требовательных молодых выводков.
Тысячи нетерпеливых молодых родителей соревнуются за лучшие места для строительства гнезд и добычи пищи для своих требовательных молодых выводков.
Исследователи наблюдали серебристых чаек, таких как эта, которые не могли стоять из-за парализованных ног. У этой и других птиц также были косые глаза, обесцвеченные зрачки и крылья, которые не складывались близко к телу.Все это симптомы дефицита тиамина. Изображение предоставлено Леннартом Балком и Пер-Оке Хегеррот (Стокгольмский университет, Стокгольм).
Но Леннарт Балк, биохимик-эколог из Стокгольмского университета, стал свидетелем совершенно иной картины, когда посетил шведские прибрежные колонии в течение 5 лет, начиная с 2004 года. Многие птицы не могли летать. Другие были полностью парализованы. Птицы также не ели и с трудом дышали. По словам Балка, тысячи птиц страдали и умирали от этой паралитической болезни.«Мы зашли в птичьи колонии и были потрясены. Вы могли видеть, что что-то действительно было не так. Это была страшная ситуация для этого времени года», — говорит он.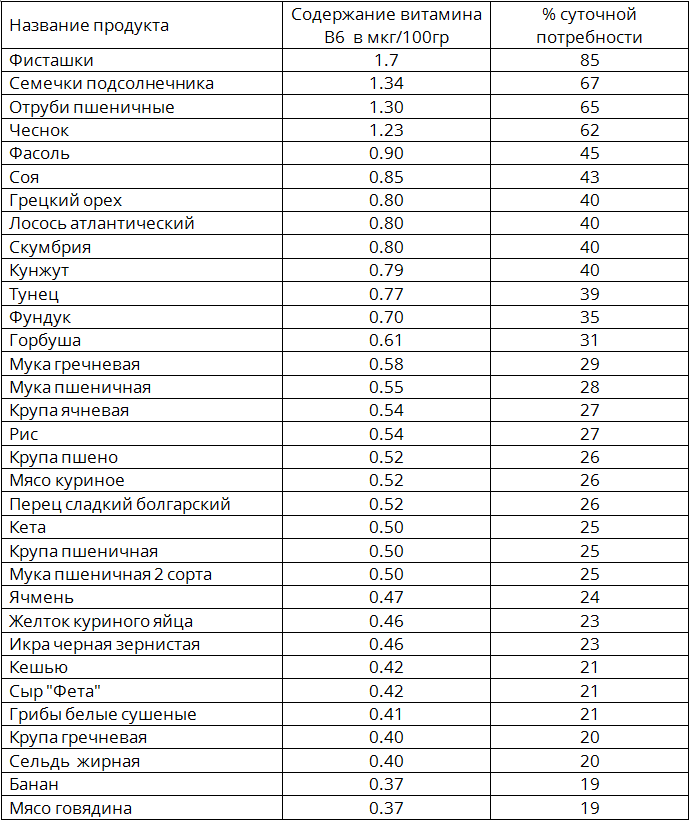
Основываясь на своей прошлой работе, документировавшей подобный кризис у нескольких видов рыб Балтийского моря, Балк предположил, что болезнь птиц была вызвана дефицитом тиамина (витамина B1). Тиамин необходим для важнейших метаболических процессов, таких как производство энергии и правильное функционирование нервной системы. Этот важный микроэлемент вырабатывается в основном растениями, в том числе фитопланктоном, бактериями и грибами; люди и животные должны получать его через пищу.
Сцены, подобные той, что в Швеции, которую в последние годы снова и снова наблюдали у различных видов в Европе и Северной Америке, заставили Балка и других исследователей забеспокоиться о том, что что-то в окружающей среде вызывает широко распространенный дефицит тиамина, что может объяснить эти специфические эпизоды, а также, возможно, крупномасштабное исчезновение популяций диких животных. «Это может быть очень серьезным источником смертности», — говорит Стивен Райли, рыбный эколог из Научного центра Великих озер Геологической службы США (USGS) в Анн-Арборе, штат Мичиган.
Исследователи в целом согласны с тем, что кризисы у морских птиц, рыб и других морских видов имеют общие недостатки тиамина. Но многое остается неизвестным. Является ли нехватка тиамина основной причиной проблемы в каждом случае? Что может быть причиной такого широко распространенного дефицита витаминов в окружающей среде? Тем не менее, поскольку случаи больных и умирающих диких животных продолжают возникать, среди исследователей, пытающихся выяснить, что происходит, возникает чувство безотлагательности. «Я не думаю, что мы согласны в нашей оценке этого, за исключением того, что это реальная проблема», — говорит Клиффорд Крафт, эколог по пресной воде из Корнельского университета в Итаке, штат Нью-Йорк.
Повторяющиеся закономерности
Будучи молодым исследователем в конце 1980-х, Балк начал изучать влияние промышленного загрязнения и эндокринных разрушителей на здоровье рыб. Но он был настолько ошеломлен, увидев влияние дефицита тиамина на рыбу, что отказался от своей предыдущей работы, чтобы вместо этого сосредоточиться на этой проблеме. В конце 1990-х Балк заметил, что несколько видов рыб в Балтийском море, в том числе атлантический лосось ( Salmo salar ), имеют проблемы с воспроизводством. Многие личинки не могли плавать прямо и были вялыми перед смертью.Когда он дополнил личинок тиамином, почти все выжили. Напротив, почти все личинки, не получавшие добавки тиамина, погибли. Балк пришел к выводу, что рыба страдает от дефицита тиамина (1).
В конце 1990-х Балк заметил, что несколько видов рыб в Балтийском море, в том числе атлантический лосось ( Salmo salar ), имеют проблемы с воспроизводством. Многие личинки не могли плавать прямо и были вялыми перед смертью.Когда он дополнил личинок тиамином, почти все выжили. Напротив, почти все личинки, не получавшие добавки тиамина, погибли. Балк пришел к выводу, что рыба страдает от дефицита тиамина (1).
Всего несколькими годами ранее исследователи начали наблюдать подобное опустошение среди рыб в Великих Лаврентийских озерах. С 1995 года исследователи, в том числе Джон Фитцсимонс, в то время биолог-рыбовед канадского правительства, а ныне на пенсии, зафиксировали дефицит тиамина, который опустошил популяции озерной форели ( Salvelinus namaycush ) и нескольких видов лосося в Великих озерах (2).Рыба не размножалась и с трудом сохраняла равновесие во время плавания. «Как будто рыба была пьяна. Они теряли равновесие и падали на бок», — говорит Фицсаймонс. Сначала исследователи думали, что виноваты загрязняющие вещества, но тесты не подтвердили это подозрение. В момент вдохновения Фицсаймонс понял, что рыбе, должно быть, не хватает питательных веществ.
В момент вдохновения Фицсаймонс понял, что рыбе, должно быть, не хватает питательных веществ.
«Сегодня с рыбой все в порядке, а на следующий день она будет лежать на боку. Это заставило меня задуматься, нет ли у них какого-то недостатка», — вспоминает он.Фитцсаймонс дополнял яйца от больных рыб тиамином и другими витаминами группы В. Он обнаружил, что менее 10% рыбьей икры, которой вводили тиамин, погибло. Другие витамины группы В не оказывали терапевтического действия. Напротив, он наблюдал более 75% смертности в контрольных образцах, которые не получали витаминов. Фитцсимонс и его коллеги пришли к выводу, что нехватка тиамина была причиной паралича и сокращения популяции рыб в Великих озерах (2, 3).
Увидев в 2004 году сходные симптомы у диких птиц, Балк сосредоточился на изучении того, как далеко зашла болезнь.В 2009 году его группа зафиксировала симптомы у серебристых чаек ( Larus argentatus ) и обыкновенного скворца ( Sturnus vulgaris ), а также у S. mollissima из участков по всей Северной Европе. Исследователи обнаружили, что концентрация тиамина в яичном желтке L. argentatus была на 41% ниже у птиц из района Балтийского моря по сравнению с птицами, взятыми из Исландии. Исследователи также заметили снижение нескольких биомаркеров активности тиамина, в том числе тиамин-зависимых ферментов, что указывает на дефицит этого витамина у некоторых птиц Балтийского моря.
mollissima из участков по всей Северной Европе. Исследователи обнаружили, что концентрация тиамина в яичном желтке L. argentatus была на 41% ниже у птиц из района Балтийского моря по сравнению с птицами, взятыми из Исландии. Исследователи также заметили снижение нескольких биомаркеров активности тиамина, в том числе тиамин-зависимых ферментов, что указывает на дефицит этого витамина у некоторых птиц Балтийского моря.
Команда Балка лечила больных птиц инъекциями тиамина и обнаружила, что все, кроме 1 из 10 парализованных взрослых особей L. argentatus , выздоровели за 2-недельный период. Ни у одной из необработанных птиц не было признаков улучшения. Кроме того, 10 молодых птиц, которых кормили раствором тиамина в первые и вторые сутки после вылупления, были бодрыми и активными, тогда как 10 молодых птиц, которых кормили физиологическим раствором, были вялыми и апатичными и начали погибать примерно через 4 дня после вылупления. Аналогичные тесты на S. vulgaris и S. mollissima также предполагают, что люди страдают от дефицита тиамина. Балк сообщил, что дефицит препятствует размножению птиц, что может способствовать сокращению местных популяций морских птиц (4).
vulgaris и S. mollissima также предполагают, что люди страдают от дефицита тиамина. Балк сообщил, что дефицит препятствует размножению птиц, что может способствовать сокращению местных популяций морских птиц (4).
Исследователи стали свидетелями того, как серебристые чайки охотятся на утят гаги, явно ослабленных из-за дефицита тиамина. Перепечатано с разрешения исх. 14.
Контрольные признаки
Такие симптомы у птиц наблюдаются не впервые.В 1880-х годах Христиан Эйкман, врач, работавший в Голландской Ост-Индии, наблюдал паралич у цыплят, отметив, что они испытывали слабость в ногах, напоминающую болезнь, известную как бери-бери, которая в то время поражала людей по всей Азии.
Эйкман связал паралич с диетой цыплят, состоящей преимущественно из вареного белого риса, который, по его мнению, должен быть токсичным; но его коллега Геррит Грейнс позже подозревал, что в результате современных процессов помола рис был лишен какого-то жизненно важного защитного вещества. В некоторых частях Азии, где рацион питания людей также в значительной степени зависел от белого риса, бери-бери поразило почти 30% населения. Эйкман получил Нобелевскую премию в 1929 году за свои наблюдения, которые помогли открыть тиамин — первый идентифицированный витамин (5). Сегодня случаи дефицита тиамина среди людей редки, потому что этот витамин добавляют в широкий спектр обработанных пищевых продуктов, таких как хлопья для завтрака. Но дефицит по-прежнему наблюдается у беженцев с плохим питанием и у алкоголиков, потому что их организм с трудом усваивает витамин.
В некоторых частях Азии, где рацион питания людей также в значительной степени зависел от белого риса, бери-бери поразило почти 30% населения. Эйкман получил Нобелевскую премию в 1929 году за свои наблюдения, которые помогли открыть тиамин — первый идентифицированный витамин (5). Сегодня случаи дефицита тиамина среди людей редки, потому что этот витамин добавляют в широкий спектр обработанных пищевых продуктов, таких как хлопья для завтрака. Но дефицит по-прежнему наблюдается у беженцев с плохим питанием и у алкоголиков, потому что их организм с трудом усваивает витамин.
Еще одной группой риска являются люди, которые едят много ферментированной рыбы, отмечает Райли, потому что рыба богата ферментом тиаминазой, расщепляющим тиамин. На самом деле, оказалось, что диета из рыбы, богатой тиаминазой, была виновником дефицита тиамина у форели и лосося Великих озер.
В 2005 году группа, в которую входил Дон Тиллитт, токсиколог-эколог из Геологической службы США, сообщила, что лосось и озерная форель питались в основном алевиками ( Alosa pseudoharengus ), инвазивными видами рыб, богатыми тиаминазой. Тиллитт и его коллеги кормили 17 самок озерной форели из инкубатория в Мичигане диетой, состоящей только из самок. Команда обнаружила, что рыба откладывала икру с общей концентрацией тиамина примерно 2,5 нмоль/г. Напротив, 13 рыб кормили только вздутиями ( Coregonus hoyi ) — рыбой-жертвой, лишенной тиаминазы — отложенной икрой, содержащей примерно 12 нмоль/г. Исследователи увидели, что почти 20% мальков погибло, когда их матерей кормили только алевками. Но все детеныши выживали, когда их матерей кормили вздутиями (6, 7).
Тиллитт и его коллеги кормили 17 самок озерной форели из инкубатория в Мичигане диетой, состоящей только из самок. Команда обнаружила, что рыба откладывала икру с общей концентрацией тиамина примерно 2,5 нмоль/г. Напротив, 13 рыб кормили только вздутиями ( Coregonus hoyi ) — рыбой-жертвой, лишенной тиаминазы — отложенной икрой, содержащей примерно 12 нмоль/г. Исследователи увидели, что почти 20% мальков погибло, когда их матерей кормили только алевками. Но все детеныши выживали, когда их матерей кормили вздутиями (6, 7).
«Дефицит тиамина почти полностью остановил размножение некоторых видов рыб в Великих озерах, что привело к резкому сокращению популяции», — говорит Тиллитт.
Верхушка айсберга?
До сих пор не появилось столь четкого объяснения для других случаев дефицита тиамина среди диких животных, задокументированных исследователями, даже несмотря на то, что число растет. В 2016 году Балк показал, что несколько других видов в Северной Европе, в том числе голубые мидии ( Mytilus sp. ) и угри ( Anguilla sp.), также страдали от дефицита. Далее он проанализировал взаимосвязь между биохимическими изменениями, вызванными дефицитом, и долгосрочными последствиями для здоровья, такими как повышенная паразитарная инфекция и нарушение роста (8).
) и угри ( Anguilla sp.), также страдали от дефицита. Далее он проанализировал взаимосвязь между биохимическими изменениями, вызванными дефицитом, и долгосрочными последствиями для здоровья, такими как повышенная паразитарная инфекция и нарушение роста (8).
«Мы обнаружили, что дефицит тиамина гораздо более распространен и серьезен, чем считалось ранее», — говорит Балк. Учитывая его масштабы, он предполагает, что повсеместный дефицит тиамина может быть, по крайней мере, частично ответственным за глобальное сокращение популяции диких животных. Например, за 60-летний период до 2010 г. популяции морских птиц во всем мире сократились примерно на 70%, а во всем мире виды исчезают в 1000 раз быстрее, чем естественные темпы исчезновения (9, 10).«Он уже наблюдал дефицит тиамина у нескольких разных типов», — говорит Фитцсаймонс из Балка. «Интересно, что происходит. Это более серьезная проблема, чем мы сначала подозревали».
В этом году в выпуске «Тенденции в экологии и эволюции» «Горизонтное сканирование новых проблем для глобальной охраны природы» видная группа исследователей в области охраны окружающей среды также отметила растущее количество свидетельств дефицита тиамина в «ряде таксономических групп» и в водах океана как « возможный фактор снижения дикой природы» (11). «Нам казалось, что это может быть важной областью, которая потребует дополнительных исследований в будущем», — говорит Уильям Дж. Сазерленд из Кембриджского университета, один из авторов.
«Нам казалось, что это может быть важной областью, которая потребует дополнительных исследований в будущем», — говорит Уильям Дж. Сазерленд из Кембриджского университета, один из авторов.
Критики, однако, говорят, что необходимы дополнительные исследования, прежде чем объявлять глобальный кризис или вовлекать нехватку тиамина. Загрязнители, такие как свинец, и такие заболевания, как ботулизм, вызываемые бактерией Clostridium botulinum , могут вызывать паралитические симптомы, сходные с симптомами дефицита тиамина. Блокируя выработку или высвобождение ацетилхолина, ботулизм вызывает потерю мышечной функции и вялый паралич ног, крыльев и шеи.В результате зараженные птицы часто тонут. Животные обычно проглатывают токсин ботулизма напрямую или поедают беспозвоночных, таких как насекомые, которые его содержат. «Связь между дефицитом тиамина и воздействием на популяционный уровень недостаточно хорошо установлена», — пишет экотоксиколог дикой природы Кристиан Сонне из Орхусского университета в Дании (12, 13). Сонне, который отказался комментировать эту статью, неоднократно критиковал статью Балка о диких птицах 2009 года.
Сонне, который отказался комментировать эту статью, неоднократно критиковал статью Балка о диких птицах 2009 года.
Тони Рок, эпизоотолог из Национального центра охраны дикой природы Геологической службы США в Мэдисоне, штат Висконсин, отмечает, что вспышки ботулизма распространены среди водоплавающих птиц, легко убивая 100 000 особей во всем мире в определенный год.По словам Рокка, в исследовании Балка не использовались тесты, необходимые для исключения других возможных причин смерти. «Вскрытие птиц не проводилось. Он просто решил, что они умерли от дефицита тиамина». Рок не оспаривает доказательства Балк о том, что животные страдали от низкого уровня тиамина, но «не соглашается с радикальным выводом» о том, что дефицит приводит к сокращению численности популяции, говорит она. Балк признает, что птицы, такие как гаги, вероятно, страдали от других болезней и последствий для здоровья, которые, как он утверждает, скорее всего, были вызваны дефицитом тиамина.
Чтобы подкрепить аргумент о том, что дефицит тиамина может быть достаточно серьезным, чтобы нанести ущерб целым популяциям, Балк и группа под руководством Торстена Мёрнера, патологоанатома из Национального ветеринарного института в Уппсале, Швеция, дополнительно изучили размножение уток гаг. Команда обнаружила, что гаги с дефицитом тиамина в Волхольме на юге Швеции откладывают меньше яиц, чем ожидалось: средний размер кладки в 16 гнездах составлял 3,8 яйца на гнездо по сравнению с нормальным средним показателем 4.6 яиц. Тесты показали, что в яйцах также не хватает тиамина.
Команда обнаружила, что гаги с дефицитом тиамина в Волхольме на юге Швеции откладывают меньше яиц, чем ожидалось: средний размер кладки в 16 гнездах составлял 3,8 яйца на гнездо по сравнению с нормальным средним показателем 4.6 яиц. Тесты показали, что в яйцах также не хватает тиамина.
В полевых условиях Мёрнер также наблюдал необычное зрелище: сотни взрослых самок в одиночестве плавали в воде. По его словам, обычно самок окружают молодые птицы. При ближайшем рассмотрении команда обнаружила, что молодые птицы выжили всего через несколько дней после вылупления. В среднем количество молодых птиц составляло всего 6% от ожидаемой численности популяции. Исследователи наблюдали за гнездами с помощью камер и видели, что яйца вылупились нормально и не были ограблены другими птицами.Вместо этого, достигнув воды, ослабленные утята стали добычей серебристых чаек (14).
«Мы обнаружили, что дефицит тиамина гораздо более распространен и серьезен, чем считалось ранее».
— Леннарт Балк
Недостаток тиамина не только привел к тому, что утки стали нести меньше яиц, говорит Мёрнер, более 90% вылупившихся утят погибли в результате нападения серебристых чаек. «Они просто лежали и смотрели на чаек, и, конечно, их съели.«Результаты Мёрнера действительно подтверждают вред, наносимый населению, — говорит Рок. «Это предполагает связь с сокращением популяции», — отмечает она, добавляя, что исследование также эффективно исключает ботулизм как причину паралича уток. «Ботулизм в этом случае был очень маловероятен».
«Они просто лежали и смотрели на чаек, и, конечно, их съели.«Результаты Мёрнера действительно подтверждают вред, наносимый населению, — говорит Рок. «Это предполагает связь с сокращением популяции», — отмечает она, добавляя, что исследование также эффективно исключает ботулизм как причину паралича уток. «Ботулизм в этом случае был очень маловероятен».
Море возможностей
Несмотря на то, что исследователи соглашаются не соглашаться с некоторыми конкретными примерами диких животных, находящихся в бедственном положении, Балк и другие исследуют, что может быть основной причиной такого широко распространенного дефицита тиамина в окружающей среде.
Балк опасается, что один распространенный фактор, такой как атмосферный загрязнитель, может истощать окружающую среду тиамином в его источниках, включая фитопланктон и бактерии, влияя на всю пищевую цепочку. Чтобы увидеть, насколько далеко заходит проблема, он теперь смотрит на наземных диких животных вверх по течению, таких как лось ( Alces alces ). Балк также исследует, может ли какой-либо из нескольких загрязнителей помешать окислению, гидролизу или синтезу тиамина.
Балк также исследует, может ли какой-либо из нескольких загрязнителей помешать окислению, гидролизу или синтезу тиамина.
Тиллит тоже забрасывает более широкую сеть, ища дефицит тиамина у водоплавающих птиц Великих озер и лосей в Миннесоте.Хотя он уверен, что причиной снижения численности рыбы в озерах были алевки, он не уверен, что может быть причиной дефицита тиамина, наблюдаемого у видов в других местах. «Если есть химическое вещество, которое каким-то образом влияет на тиамин, это может быть чрезвычайно опасно», — говорит он. «Для нас очень важно узнать об этом больше».
Но исследователям не нужно ссылаться на загрязняющие вещества, чтобы объяснить дефицит тиамина, говорит Серджио Санудо-Вильгельми, эколог-биогеохимик из Университета Южной Калифорнии в Лос-Анджелесе.Проблема может быть связана с дисбалансом фитопланктона и бактерий, которые являются основными производителями тиамина и других витаминов группы В (15).
Саньюдо-Вильгельми измерил очень низкий уровень витаминов группы В, включая тиамин, в прибрежных водах Калифорнии. Другие исследователи оценили аналогичный дефицит в некоторых районах открытого океана (16). По его словам, потепление воды из-за изменения климата может объяснить нехватку витаминов в морской воде. Более высокие температуры ускоряют рост бактерий, заставляя микробы потреблять больше витаминов группы В, чем они производят, — поглощая витамины до того, как фитопланктон сможет получить свою долю.
Другие исследователи оценили аналогичный дефицит в некоторых районах открытого океана (16). По его словам, потепление воды из-за изменения климата может объяснить нехватку витаминов в морской воде. Более высокие температуры ускоряют рост бактерий, заставляя микробы потреблять больше витаминов группы В, чем они производят, — поглощая витамины до того, как фитопланктон сможет получить свою долю.
Саньюдо-Вильгельми предполагает, что несколько иной дисбаланс мог вызвать дефицит тиамина в районе Балтийского моря, где, вероятно, играют роль азотные и фосфорные загрязнения. Большое цветение цианобактерий — токсичных сине-зеленых водорослей — обычны в Балтийском море летом из-за эвтрофикации. Работа исследователей из Университета Линнея в Швеции показала, что зоопланктону — крошечным водным животным, питающимся фитопланктоном, — не хватало тиамина во время такого цветения (17).В результате витамин больше не передается по пищевой цепочке к мелкой рыбе, питающейся зоопланктоном, или к крупным хищникам, питающимся этой рыбой, говорит один из авторов исследования, акваэколог Сэмюэл Хайландер. Санудо-Вильгельми говорит, что растущее число цветков токсичных цианобактерий, происходящих по всему миру, может вызвать аналогичный дефицит тиамина в других местах, что предполагает еще один потенциальный путь распространения проблемы.
Санудо-Вильгельми говорит, что растущее число цветков токсичных цианобактерий, происходящих по всему миру, может вызвать аналогичный дефицит тиамина в других местах, что предполагает еще один потенциальный путь распространения проблемы.
Но Санюдо-Вильгельми говорит, что еще слишком рано говорить о том, что нехватка витаминов в океане, которую он задокументировал вдоль побережья Тихого океана, происходит где-то еще.Имеются ограниченные данные о концентрациях растворенных витаминов в разных географических районах и в разные сезоны, поэтому у исследователей нет доказательств общих тенденций в уровнях содержания витаминов в окружающей среде.
Если причиной дефицита тиамина в разных условиях являются различные виды дисбаланса, тайну тиамина можно разбить на более управляемые части. Например, нехватку рыбы в Великих озерах удалось решить путем добавления тиамина в воду в рыбоводных хозяйствах; затем искусственно выращенную рыбу выпускали в озера для увеличения популяции.В конце концов, количество бабочек также начало снижаться, и естественное воспроизводство форели и лосося в озерах начало восстанавливаться (18). И если цветение цианобактерий играет определенную роль, усилия по сдерживанию сельскохозяйственного загрязнения могут помочь предотвратить распространение дефицита по спирали вверх по пищевой цепочке.
И если цветение цианобактерий играет определенную роль, усилия по сдерживанию сельскохозяйственного загрязнения могут помочь предотвратить распространение дефицита по спирали вверх по пищевой цепочке.
Это кажется очевидным: универсальное решение более широкой проблемы маловероятно. По словам Балка, добавки с тиамином не являются реальным решением для улучшения целых экосистем.
Независимо от того, является ли виновником один тиамин или нет, Балк и другие намерены собрать больше улик в надежде разгадать тайну, имеющую серьезные последствия для многих видов и экосистем.«Самое главное, — говорит Балк, — это найти причину».
9 Преимущества тиамина (витамина B1) + источники, дозировка
Тиамин, или витамин B1, является важным питательным веществом, необходимым организму. Он имеет много преимуществ для здоровья, от защиты мозга и сердца до укрепления иммунной системы.
Что такое витамин B1 (тиамин)? Тиамин, также известный как витамин B1, необходим для каждой ткани организма. Он является кофактором ферментативных реакций в скелетных мышцах, сердце, печени, почках и головном мозге [1].
Он является кофактором ферментативных реакций в скелетных мышцах, сердце, печени, почках и головном мозге [1].
Поступивший внутрь Витамин B1 из пищевых продуктов и пищевых добавок всасывается в тонком кишечнике посредством активного транспорта в пищевых дозах и путем пассивной диффузии в фармакологических дозах [1].
Большая часть пищевого витамина B1 находится в фосфорилированных формах, и фосфатазы кишечника гидролизуют их, освобождая тиамин до того, как витамин всасывается. Люди хранят тиамин в основном в печени, но в очень небольших количествах [2].
Снимок- Необходим для обмена веществ
- Повышает иммунную систему
- Поддерживает функцию мозга
- Помогает пищеварению
- Защищает сердце
Организму необходим витамин B1 для выработки АТФ, основной молекулы, несущей энергию в организме.
Тиамин помогает в преобразовании углеводов в глюкозу, которая является предпочтительным источником энергии, из которой организм расходует энергию, чтобы обеспечить бесперебойную работу метаболизма.Он также помогает расщеплять белки и жиры [3].
Тиамин особенно необходим для системы ферментативных реакций, называемой пируватдегидрогеназой, которая участвует в метаболизме сахаров, которые мы едим [4].
Метаболизм сахара
Тиамин (в виде тиаминдифосфата, основной активной формы витамина) необходим для метаболизма глюкозы [5].
Доля людей с диабетом 1 или 2 типа, у которых низкий уровень тиамина, колеблется от 17% до 79%. Исследования показали, что увеличение потребления витамина B1 снижает тяжесть симптомов, связанных с диабетом на ранней стадии [6].
2) Иммунитет Как и другие витамины группы В, витамин В1 иногда называют «антистрессовым» витамином, поскольку он может укреплять иммунную систему и улучшать способность организма противостоять стрессовым условиям [7].
Введение крысам блокатора витамина B1 вызывало значительное снижение функции иммунной системы [7].
3) МозгПредыдущие исследования сообщали о низком уровне дисфункции тиамина и пируватдегидрогеназы у пациентов с атаксией, состоянием, которое вызывает потерю движения.Длительное лечение показало значительные улучшения [8].
Витамин B1, по-видимому, помогает в развитии миелиновой оболочки , оболочки, которая оборачивается вокруг нервов, чтобы защитить их от повреждения и гибели [9].
В головном мозге он требуется как нервным клеткам, так и другим опорным клеткам нервной системы [10].
Вскрытие показало, что тиаминзависимые ферменты снижают активность в мозге людей с болезнью Альцгеймера [11].
Дополнительный тиамин и мозг
Высокие дозы тиамина уменьшали усталость у пациентов после инсульта [12].
Некоторые исследователи подозревают, что терапия витамином B1 может оказывать благоприятное влияние на нейродегенеративные заболевания, такие как болезнь Паркинсона, болезнь Альцгеймера и Хантингтона [13].
Необходимо провести дополнительные исследования, прежде чем можно будет сделать какие-либо выводы о том, может ли он помочь людям с нейродегенеративными заболеваниями.
4) Здоровье сердечно-сосудистой системыТиамин жизненно важен для функции сердечно-сосудистой системы, а дефицит тиамина может вызвать застойную сердечную недостаточность [5].
Дополнительный тиамин и сердце
В обзоре 20 клинических исследований добавление тиамина улучшало сердечную функцию у людей с сердечной недостаточностью [14].
По сравнению с плацебо, прием тиамина в 2 рандомизированных двойных слепых исследованиях привел к значительному улучшению фракции выброса левого желудочка [15].
5) Катаракта Недавние исследования показывают, что тиамин может снизить риск развития катаракты . Эти исследования показывают, что люди, которые потребляют много белка вместе с витаминами A, B1, B2 и B3 (или ниацином) в своем рационе, менее склонны к развитию катаракты. Получение достаточного количества витаминов С, Е и витаминов группы В дополнительно защищает хрусталик глаза (16).
Эти исследования показывают, что люди, которые потребляют много белка вместе с витаминами A, B1, B2 и B3 (или ниацином) в своем рационе, менее склонны к развитию катаракты. Получение достаточного количества витаминов С, Е и витаминов группы В дополнительно защищает хрусталик глаза (16).
Тиамин также необходим для правильного функционирования пищеварительной системы . Витамин B1 помогает регулировать выработку соляной кислоты, которая необходима для поддержания правильной пищеварительной функции [17].
Дефицит тиамина
Синдром Вернике-Корсакова
Дефицит тиамина является установленной причиной синдрома Вернике-Корсакова (WKS), связанного с алкоголем неврологического расстройства. Употребление алкоголя может повредить мозг посредством многочисленных механизмов; один из этих механизмов включает снижение доступности витамина B1 для мозга вследствие продолжительного употребления алкоголя [10].
Авитаминоз
Дефицит тиамина вызывает бери-бери. Некоторые из симптомов включают отек, покалывание или жжение в руках и ногах, а также затрудненное дыхание из-за жидкости в легких [18].
Люди в развитых странах обычно не заболевают авитаминозом, потому что основные продукты питания, такие как крупы и хлеб, обогащены витамином B1 [5].
Источники тиамина| Продукты питания | Сервировка | Тиамин (мг) |
| Чечевица (вареная, вареная) | ½ стакана | 0,17 |
| Зеленый горошек (вареный, вареный) | ½ стакана | 0.21 |
| Длиннозерный коричневый рис (вареный) | 1 чашка | 0,19 |
| Длиннозерный белый рис, обогащенный (вареный) | 1 чашка | 0,26 |
| Длиннозерный белый рис, необогащенный (вареный) | 1 чашка | 0,04 |
| Цельнозерновой хлеб | 1 ломтик | 0,10 |
| Белый хлеб (обогащенный) | 1 ломтик | 0,23 |
| Обогащенные сухие завтраки (пшеничные, воздушные) | 1 чашка | 0. 31 31 |
| Сухие завтраки из зародышей пшеницы (поджаренные, простые) | 1 чашка | 1,88 |
| Свинина постная (корейка, вырезка, вареная, жареная) | 3 унции* | 0,81 |
| Пекан | 1 унция | 0,19 |
| Шпинат (приготовленный, вареный) | ½ стакана | 0,09 |
| Оранжевый | 1 фрукт | 0,11 |
| Дыня | ½ фрукта | 0.11 |
| Молоко | 1 чашка | 0,10 |
| Яйцо (вареное, сваренное вкрутую) | 1 большой | 0,03 |
| *3 унции мяса — это порция размером с колоду карт [11] | ||
Нагревание продуктов может снизить содержание в них тиамина . Например, пастеризация снижает содержание тиамина в молоке на 20 % [2].